
Carpocladus fertilis Vervoort & Watson, 2003
|
publication ID |
https://doi.org/10.11646/zootaxa.5082.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:D868FB39-82D3-4413-B0FD-33808CE1CCB3 |
|
DOI |
https://doi.org/10.5281/zenodo.5792785 |
|
persistent identifier |
https://treatment.plazi.org/id/038D87B5-A136-A626-FF56-0D0211C6FD11 |
|
treatment provided by |
Plazi |
|
scientific name |
Carpocladus fertilis Vervoort & Watson, 2003 |
| status |
|
Carpocladus fertilis Vervoort & Watson, 2003 View in CoL
Figs 1–6 View FIGURE 1 View FIGURE 2 View FIGURE3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 1 View TABLE 1
Carpocladus fertilis Vervoort & Watson, 2003: 282 View in CoL , fig. 67.
Material examined. MNHN-IK-2015-3028, KANADEEP 2, Stn. DW 5107, off southern New Caledonia, Mount Munida , 22°59.9’ S, 168°14.4’ E, 30 Sep 2019, 1151– 1274 m, a ca. 5 cm high cormoid bearing two phylactocarps with female gonothecae. GoogleMaps —MNHN-IK-2015-3026, KANADEEP 2, Stn. CP 5095, off southern New Caledonia, Capucine Bassin, 23°38’ S, 167°53.1’ E, 29 Sep 2019, 1087– 1081 m, a ca. 4 cm high cormoid bearing three phylactocarps with male gonothecae GoogleMaps .
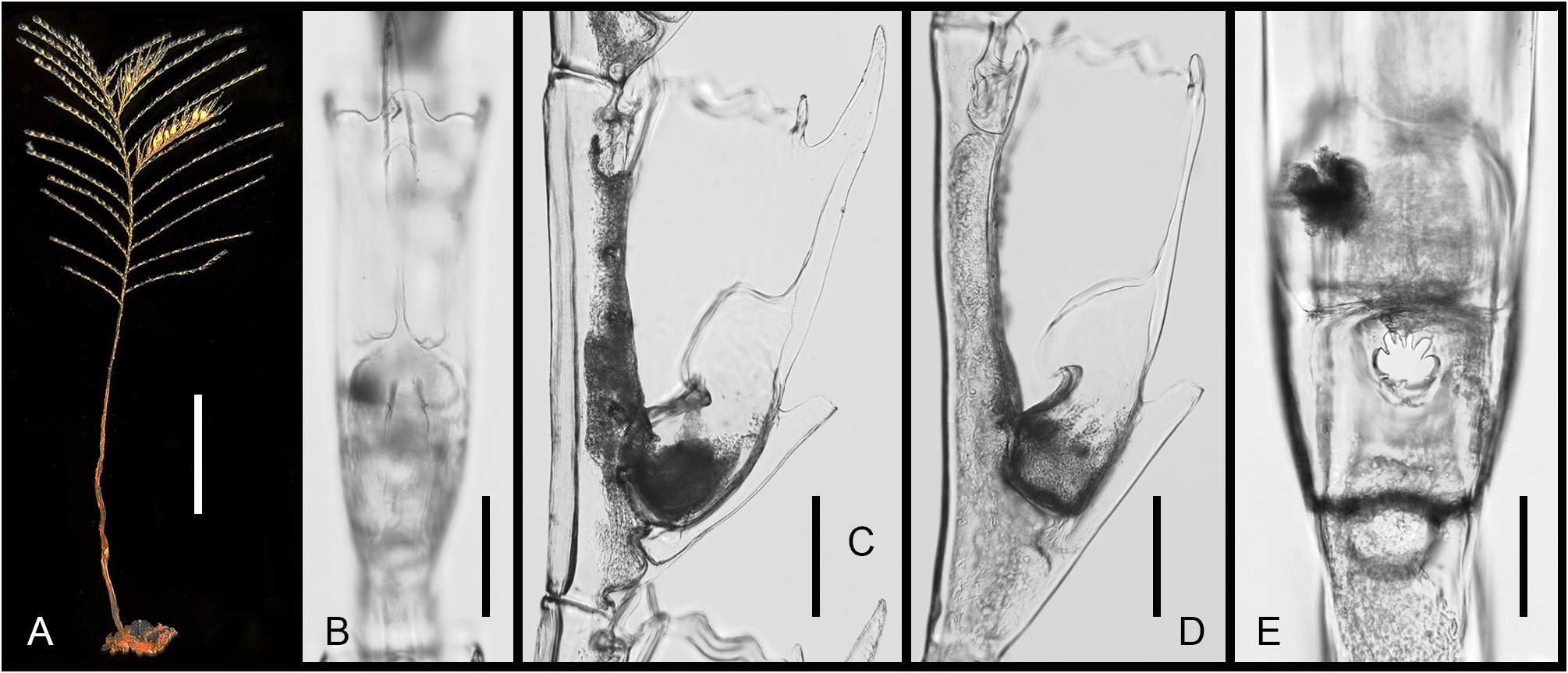
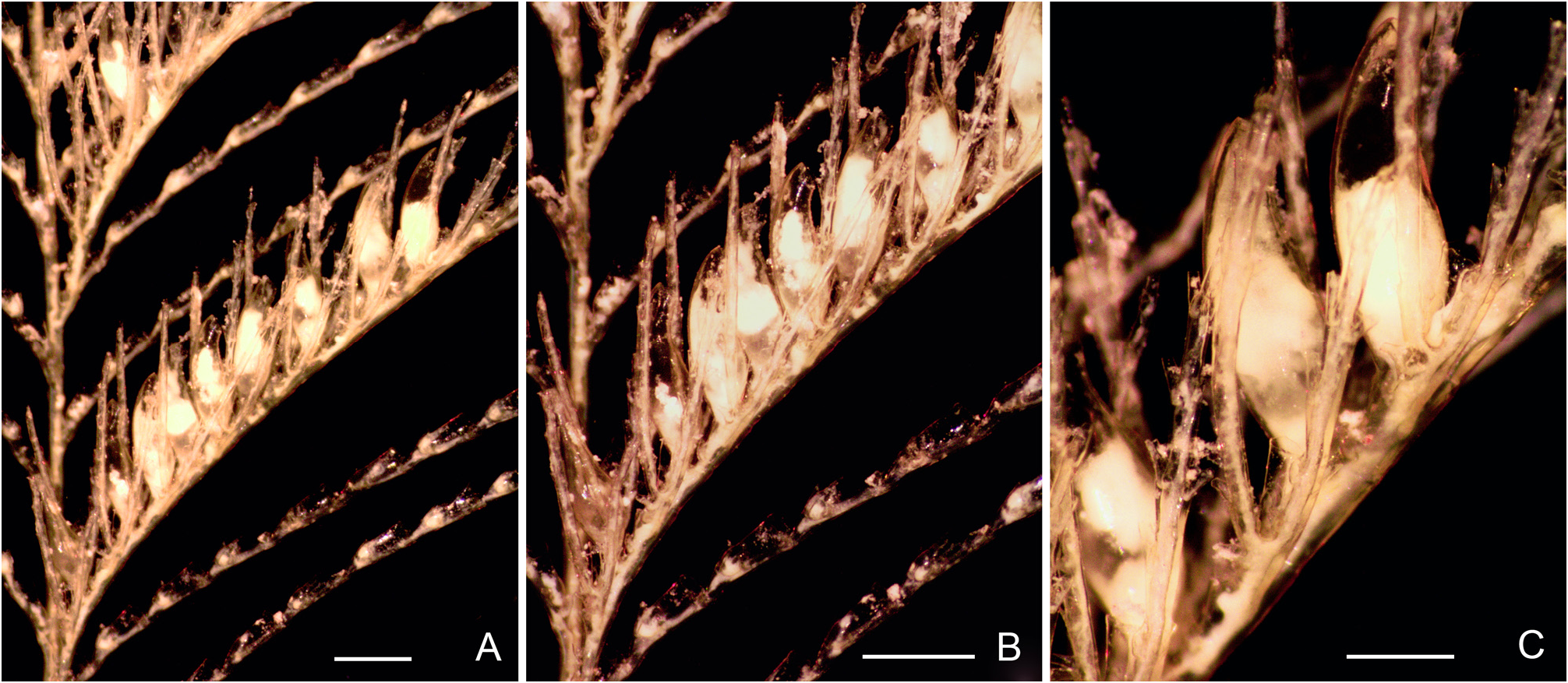
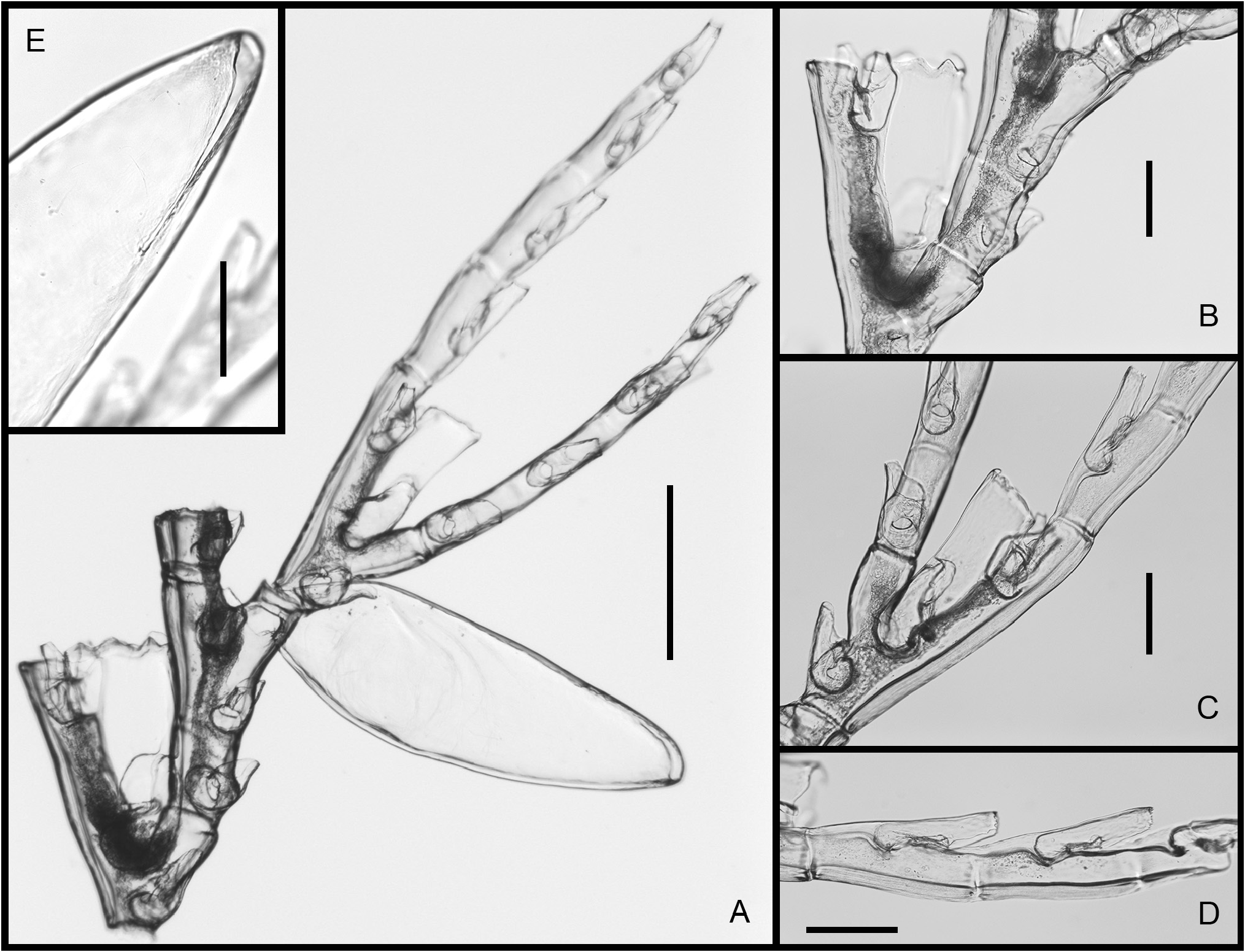
Description. Colonies up to 5 cm high, arising from rhizoid stolons firmly attached to pebbles ( Fig. 1A View FIGURE 1 ). Stems unbranched, lightly fascicled proximally, unable to support themselves when out of liquid; divided into a proximal, acladiate part of varied length, and a generally longer, distal part bearing alternate hydrocladia; both parts separated by up to five prosegments delimited by deeply-incised hinge-joints, each bearing one or two frontal nematothecae; hinge-joint region distinctly flattened laterally, allowing the colony to swing freely in the water currents; upper part of stem regularly-divided into internodes by means of oblique constrictions of the perisarc; internodes moderatelylong, each with a lateral apophysis in upper half, an axillar nematotheca, and a centrally-placed nematotheca in the lower half ( Fig. 2A View FIGURE 2 ); cauline nematothecae scoop-shaped: axillar ones narrower, lower ones comparatively broader ( Fig. 2B–D View FIGURE 2 ); apophyses short, alternate, given off at an angle of about 60° with the long axis of the internodes, and slightly frontally ( Fig. 2B View FIGURE 2 ); apophyses separated by the corresponding cladia by a slightly oblique node. Cladia unbranched, up to 14 mm long, arching gracefully downward distally; divided into up to 14 internodes by regular, transverse nodes; internodes long, each accommodating a hydrotheca occupying upper 4/5 of their length ( Fig. 2E, H View FIGURE 2 ). Hydrothecae deep, slightly inverted-conical, laterally-flattened, fully adnate, aperture ovoid, distal, slightly tilted forwards ( Figs 1B–D View FIGURE 1 ; 2E–H View FIGURE 2 ); an internal, transverse, upwardly-directed septum projecting into the lower fourth of the lumen, distal edge backwardly-rolled ( Fig. 2K View FIGURE 2 ); above, two lateral, sigmoid septa meeting frontally divide the hydrothecal lumen into a lower third and an upper two-thirds, leaving a V-shaped passage for the hydranth ( Fig. 2K, L View FIGURE 2 ); several short perisarc plugs project into the internode behind the adaxial hydrothecal wall: one prolongs the transverse septum in the opposite direction, another one is given off at junction between the hydrothecal base and the adaxial wall ( Fig. 2K View FIGURE 2 ), and two others are occasionally present behind the adaxial wall (not shown); an additional perisarc plug is to be found proximally on the adaxial side of the cladial internode ( Fig. 2K View FIGURE 2 ); below the intrathecal transverse septum, there is a round hydropore surrounded by whorl of spines ( Fig. 1E View FIGURE 1 ); hydrothecal rim with a broad, median, rounded cusp with thickened perisarc ( Fig. 1B View FIGURE 1 ), and four lateral pairs of triangular cusps, those of the first pair distinctly blunted, and those of the fourth pair partly fused to the lateral nematothecae ( Figs 1C, D View FIGURE 1 ; 2I View FIGURE 2 ); adaxially, hydrothecal rim broadly scooped; most hydrothecae are provided with a slender, horn-like, upwardlycurved, hollow perisarc projection ( Figs 1B, C View FIGURE 1 ; 2E, I View FIGURE 2 ) of varied development (though generally unmistakable) and prolonged proximally into a short, longitudinal, median carina. Three nematothecae associated to each hydrotheca: a mesial one and a pair of laterals; mesial nematotheca originating slightly below the hydrothecal base, long, tubular, adnate for 4/5 of its length, aperture gutter-shaped, rim distinctly crenulate ( Fig. 2K View FIGURE 2 ); lateral nematothecae slightly surpassing the hydrothecal rim, monothalamic, urn-shaped, with deep, adaxial emargination, rim crenulate ( Fig. 2I View FIGURE 2 ). Phylactocarps borne on the proximal-most cormidia ( Fig. 3A View FIGURE3 ), and given off slightly in front of the cormoid ( Fig. 1A View FIGURE 1 ); up to 9 mm long; first cormidium with a short, lateral, athecate apophysis inserted below the hydrothecal base, thus displacing the mesial nematotheca towards the proximal end of the internode, and forcing it to adopt the same shape as the cauline nematothecae ( Figs 4A View FIGURE 4 ; 6A View FIGURE 6 ); phylactocarp composed of a proximal-most, short internode bearing a median nematotheca ( Figs 4A View FIGURE 4 ; 5A, B View FIGURE 5 ; 6A View FIGURE 6 ), followed by a regular succession of up to 16 geniculate internodes forming its rachis ( Fig. 4B View FIGURE 4 ); rachial internodes geniculate, moderately-long, composed of a lateral apophysis supporting a costa (apophyses alternate), an axillar nematotheca, and another nematotheca proximally ( Fig. 6A View FIGURE 6 ); nematothecae scoop-shaped; costae widely-spaced ( Fig. 4B View FIGURE 4 ), phylactocarp open ( Fig. 3 View FIGURE3 ); costae bifid ( Figs 3C View FIGURE3 ; 5A View FIGURE 5 ; 6A View FIGURE 6 ), with a basal, smaller hydrotheca ( Fig. 5C View FIGURE 5 ) whose modified mesial nematotheca is displaced proximally; hydrothecate internode prolonged distally by up to six short, collinear internodes, each provided with a single or a pair of elongate nematothecae set laterally; between the mesial nematotheca and the hydrothecal base, a slightly laterally-projecting apophysis supporting the second branch of the costa; second branch with up to four nematothecate internodes ( Fig. 6A View FIGURE 6 ); nematothecae half-adnate, gutter-shaped distally, with crenulate rim ( Fig. 5D View FIGURE 5 ). Gonothecae borne on inconspicuous swellings of the rachial apophyses supporting the costae, the latter arching over the gonothecae; gonothecae fusiform, sexually dimorphic, female ( Figs 4C View FIGURE 4 ; 6D View FIGURE 6 ) comparatively longer and curving away distally ( Fig. 6C View FIGURE 6 ), and with wider distal apertures than the male ones (compare Fig. 4C and B View FIGURE 4 , and 6D and B View FIGURE 6 , respectively); apertures set distally, transverse, circular in shape, prolonged downward as long slits ( Figs 4B, C View FIGURE 4 ; 5E View FIGURE 5 ; 6B, D View FIGURE 6 ). Coenosarc poorly-preserved, the number of tentacles in hydranths and of oocytes in the female gonothecae could not be ascertained. Perisarc of colony straw colored.
Remarks. Cormoids in the present material are comparatively smaller ( 4–5 cm high) than in the holotype colony ( 12 cm high). Consequently, their stems are less fascicled, and their cladia shorter (up to 14 mm vs. 25–30 mm long in the holotype), comprising lesser cormidia (14 vs. up to 35) ( Vervoort & Watson 2003). Reportedly, up to 5 internal, incomplete, annular ridges can be found in the internode behind the hydrotheca. Unlike in the present material, the pedicel of the phylactocarp could be quite long, and composed of up to 12 nematothecate internodes, as noted in the holotype. Not stated in the original account, the nematothecae of this species have distinctly crenulated rims.
Prior to this study, only the male gonothecae have been described ( Vervoort & Watson 2003). However, one of the cormoids examined here bears phylactocarps with female gonothecae. Similar in shape to their male counterparts, but larger, they nevertheless possess comparatively broader apertures apically (compare Fig. 6D and 6B View FIGURE 6 , respectively).
The genus Carpocladus was created to accommodate Cladocarpus -like hydroids producing rather complex phylactocarps whose lateral, alternate costae are bifid, each being provided with an axillar hydrotheca between two flanking nematothecate ramuli. In Vervoort & Watson’s (2003) view, their genus showed closer affinities with Cladocarpoides Bogle, 1984 , in which the exceedingly long phylactocarps have costae also provided with a hydrotheca (ending in a “nematophorous spine”), in front of which (between the proximally-displaced mesial nematotheca and the hydrothecal base) arises an antler-shaped, “nematophorous branchlet” or “phylactogonium” ( Bogle 1984). Strikingly, the relationships between Carpocladus and Wanglaophenia Vervoort & Watson, 2003 , a hydroid genus forming nearly identical phylactocarps, were not discussed.
Given the lack of genetic material, not only of C. fertilis , but also of many Cladocarpus species and members of Cladocarpus -like genera, and the unpredictable relationships between the few species for which genetic data are already available ( Moura et al. 2018), it is impossible for the time being to ascertain the validity of the genus Carpocladus , as well as its phylogenetic position within the family Aglaopheniidae .
Distribution. Tasman Sea ( Vervoort & Watson 2003), off southern New Caledonia (present study). Occurs at depths between 979–1274 m [ Vervoort & Watson (2003) and present study, respectively].



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Hydroidolina |
|
Order |
|
|
Family |
|
|
Genus |
Carpocladus fertilis Vervoort & Watson, 2003
| Galea, Horia R. 2021 |
Carpocladus fertilis
| Vervoort, W. & Watson, J. E. 2003: 282 |