
Paraisanthus fabiani, Häussermann, Verena & Försterra, Günter, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.184465 |
|
DOI |
https://doi.org/10.5281/zenodo.5688855 |
|
persistent identifier |
https://treatment.plazi.org/id/03829541-FFB8-FFF0-29A2-FCE77550FB4D |
|
treatment provided by |
Plazi |
|
scientific name |
Paraisanthus fabiani |
| status |
sp. nov. |
Paraisanthus fabiani View in CoL new species
External anatomy ( Fig. 2 View FIGURE 2 ). Size in vivo, measured in aquaria: oral disc 7–12 mm diameter, column to 10 mm diameter, pedal disc to 19 mm diameter, 13–20 mm long, tentacles about half as long as diameter of oral disc. Size preserved: oral disc to 8 mm diameter, pedal disc to 19 mm diameter, column to 16 mm long, tentacles to 5 mm long.
Colour (Figs. 3A–G). Oral disc red, orange, yellowish, pale rose, pink, light brown or ochre; uniformly coloured or with (often 12) reddish-brown radial lines visible on endocoels, or with 12-lobed, petal-like white and yellow pattern ( Fig. 2 View FIGURE 2 B). Mesenterial insertions visible through oral disc. Actinopharynx yellow to light brown. Tentacles slightly transparent, rose to red or pink, yellowish, light brown or ochre; uniformly coloured ( Figs. 2 View FIGURE 2 D–G) or with 3–6 brown transverse bands at inner and outer or only inner side ( Figs. 2 View FIGURE 2 A–C). Column rose-coloured, reddish, yellowish, orange to ochre, or light brown; uniformly coloured or distalmost 1 / 4–1 / 5 more intensely coloured (often brown; Fig. 2 View FIGURE 2 D) with a short transition to paler proximal portion of column; colour generally continuously fading towards pedal disc. Pedal disc coloured as column, not transparent. Preserved specimens whitish to brown.
Oral disc and tentacles. Between 66 and 110 conical tentacles, hexamerously arranged in 5–6 cycles, last cycle generally not complete, length about half diameter of oral disc, inner longer than outer, situated on outer third to half of oral disc, innermost 12 (two cycles) in many animals slightly more central ( Fig. 2 View FIGURE 2 A), more intensely coloured and directed upward. Oral disc circular, mesenterial insertions in many specimens visible as darker lines ( Figs. 2 View FIGURE 2 A,C,D,E). Mouth opening central, slightly oval, slightly elevated in many specimens.
Column. In situ higher than broad, after sampling broader than high, proximally broader than distally; smooth, often with loose ring of mucus and dirt in proximal part ( Figs. 2 View FIGURE 2 C,D,G). Small fosse. Column can completely cover tentacles when retracted.
Pedal disc. More or less circular, generally wider than column and oral disc, limbus slightly lobed.
Internal anatomy. In most parts of column, 24 mesenteries hexamerously arranged in four cycles, first cycle (six pairs) including directives fertile macrocnemes with strong circumscript retractors, second and third cycle (six plus 12 pairs) sterile microcnemes without retractors, fourth cycle incomplete, pairs of extremely small microcnemes only just below margin, unequal proximal extension of mesenteries of a pair. Mesenteries of second cycle wider than those of third cycle. More tentacles than mesenteries in mid-column and at base. Actinopharynx deeply furrowed, with two distinct siphonoglyphs, about half length of column; two pairs of FIGURE 3. Histological sections of Paraisanthus fabiani : A, macrocneme with retractor and parietobasilar muscle; B, transverse section of the upper column; C, transverse section of the lower column; D, longitudinal section of the upper column with sphincter; E, cross section through tentacle; F, longitudinal section of the pedal disc with basilar muscles. 1st to 3rd cycle of mesenteries I, II, III, basilar muscles bm, directives d, ectoderm ec, endoderm en, ectodermal longitudinal muscles of tentacle et, mesogloea m, macrocnemes ma, filaments mf, microcnemes mi, parietobasilar muscles pb, actinopharynx ph, retractor muscles r, sperms s, siphonoglyph si, sphincter sp.
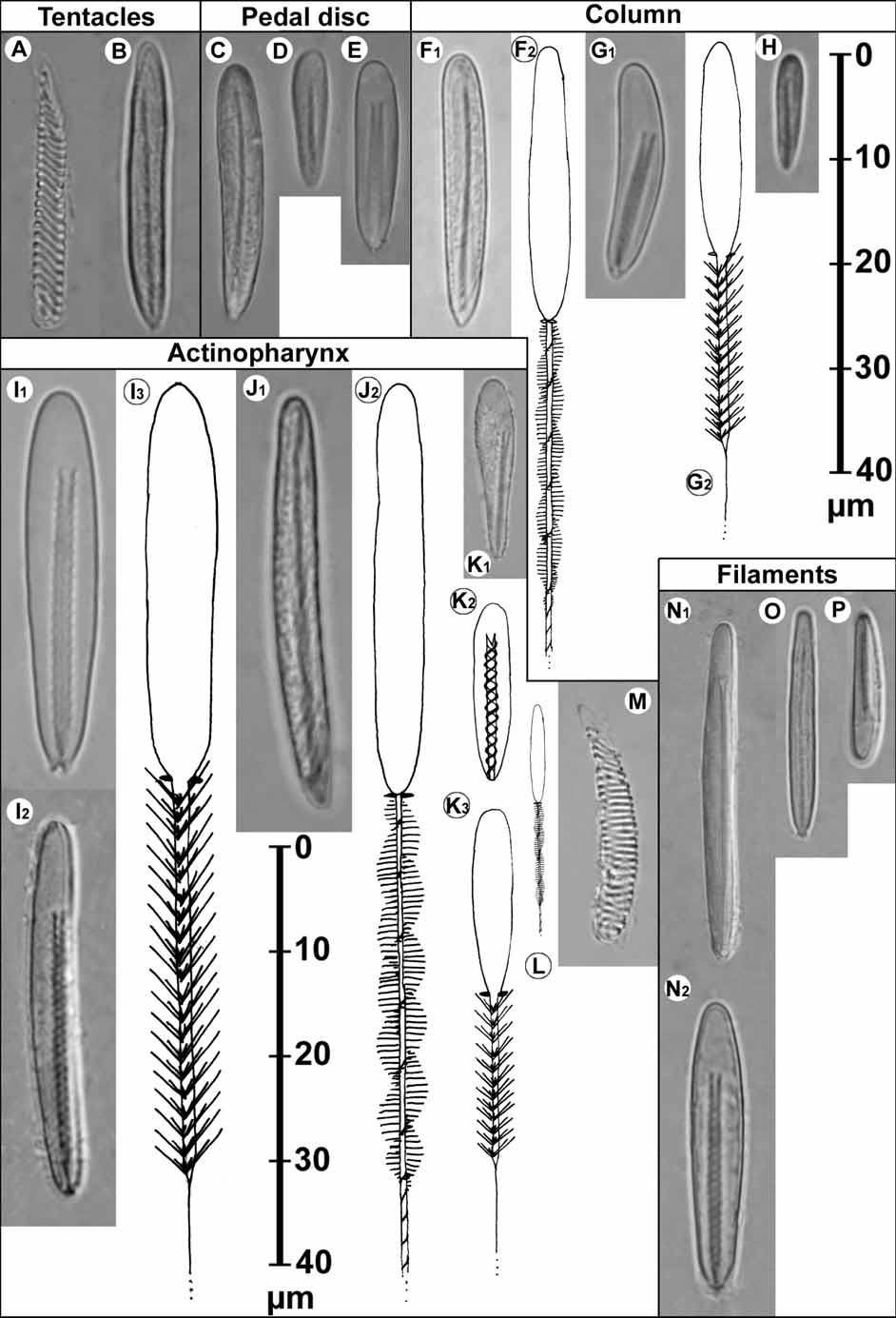
TABLE 2. Size and distribution of cnidae of Paraisanthus fabiani n. sp. (ZSM 20070247/1, letters refer to Fig. 4 View FIGURE 4 ), in each tissue in order of abundance: s: sporadic, f: few, c: common and v: very common. “m l“ and “m w” are the means, “d l” and “d w” are the standard deviations (all in µm), “t” is the apparent total number of turns of spine-rows on the shaft, “#” is the number of capsules measured, “p” is the proportion of specimens having this cnida type. Exceptional sizes in parentheses. Spirocysts (A,M), basitrichs (B,C,D,F,H,J,L,O), microbasic p-mastigophores (E,N,P), microbasic pmastigophores B1 (G,I,K).
directives. Oral and marginal stomata; marginal stomata circular, in the centre of stronger mesenteries (stomata not always visible in smaller specimens). Sexes separate, no signs of asexual reproduction. Five of the sectioned specimens with reproductive tissue, collected in February and March, four male (ZSM 20070246, ZSM 20070247/1, ZSM 20070249, USNM 1101612), one female (ZSM 20051690). No zooxanthellae.
Sphincter mesogloeal, strong, nearly entire width of mesogloea (Fig. 3D), restricted to uppermost part (~ 1/8–1/10) of column. Macrocneme retractors strong, strongly restricted to circumscribed (Figs. 3A–C). Parietobasilar muscles distinct on perfect mesenteries (Figs. 3A–C); basilar muscles distinct (Fig. 3F). Longitudinal muscles of tentacles (Fig. 3E) and radial muscles of oral disc ectodermal. Endodermal circular muscles of column well marked, weaker at sphincter level (Fig. 3D).
Cnidom. Spirocysts, basitrichs, microbasic p-mastigophores B ( Fig. 4 View FIGURE 4 ).
Cnidae of eight specimens were examined.
Etymology. The species is dedicated to our son Fabian who was born on May 16th 2007 some days after the submission of this paper.
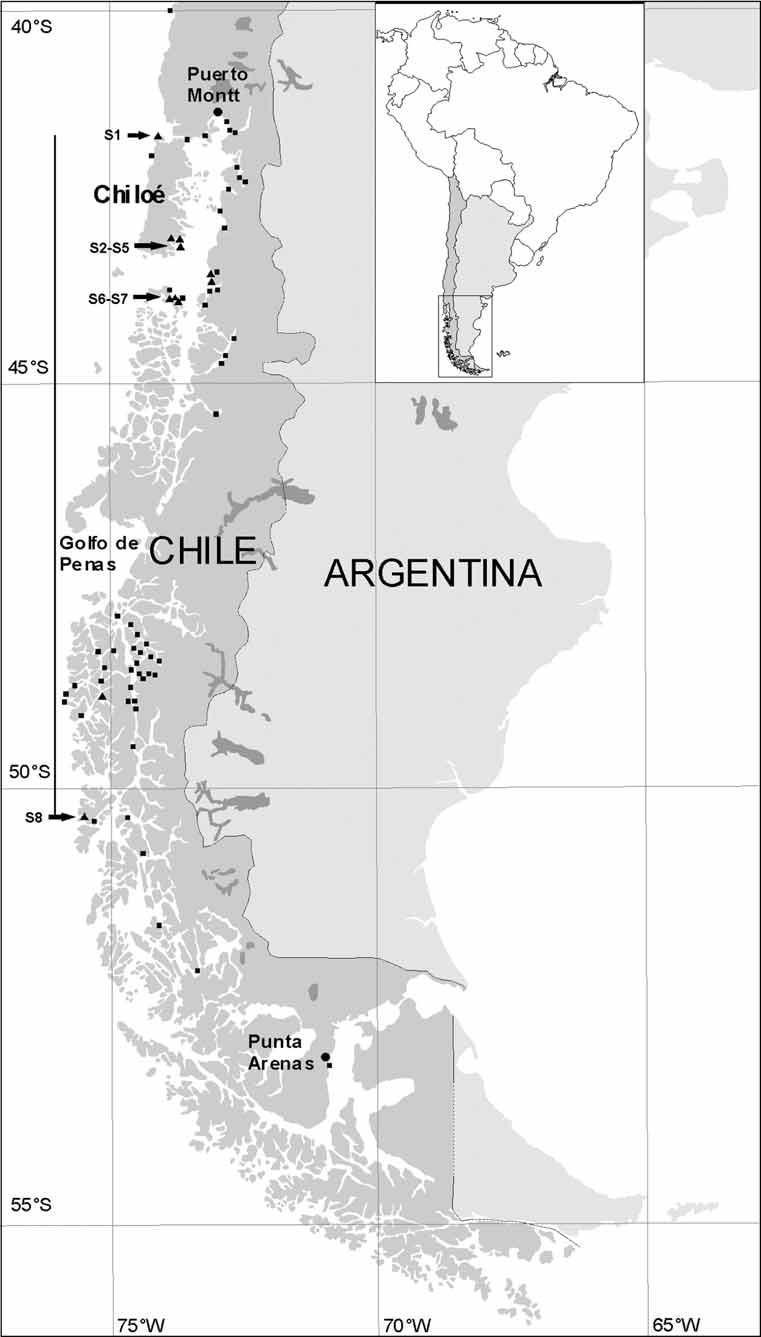
Habitat, distribution, and zoogeography. Shallow subtidal to at least 30 m in less protected channels and at exposed islands of Chilean fjord region between Faro Corona, Chiloé Island (41°47’02.0’’S; 73°52’58.8’’W) and Archipelago Madre de Dios (50°20’23.1"S; 75°22’39.2"W) ( Fig. 1 View FIGURE 1 ). Not present along exposed coast north of fjord region between Faro Corona, North Chiloé Island, and Arica, North Chile (41°47’S to about 15°S), or in southern third of fjord region between Archipelago Madre de Dios and Straits of Magellan (50°20’S to about 53°S) ( Fig. 1 View FIGURE 1 ); neither present in protected fjords and channels with superficial fresh water layer. Thus, it is present in both the Northern and the Central Patagonian Zone (see Pickard 1971; Häussermann 2006), which are separated by Peninsula Taitao and Golfo de Penas ( Fig. 1 View FIGURE 1 ), hypothesized by Lancellotti and Vasquez (2000) and Häussermann and Försterra (2005) to be zoogeographic barriers, but not in the Southern Patagonian Zone south of the Straits of Magellan.
Natural history and field notes. Not abundant at any site, but if present, generally several clustered specimens found ( Fig. 5 View FIGURE 5 ). Pedal disc and column generally hidden in holes or crevices of rocky substrate, between stones or between polychaete tubes into which entire animal retracts when disturbed ( Fig. 5 View FIGURE 5 ). Tentacles of neighbouring specimens regularly in contact with each other. Specimens observed in same habitat with the sea anemones Halcurias pilatus McMurrich, 1893 , Phellia exlex McMurrich, 1904 , Sagartiidae sp. ( Fig. 5 View FIGURE 5 C), and a corallimorpharian ( Corynactis sp.); in Central Patagonian Zone also with the sea anemone Metridium senile lobatum Carlgren, 1899 ( Riemann-Zürneck 1975) .
Difficult to collect without injury due to cryptic microhabitats; ideally collected with hammer and chisel together with substrate. Specimens relatively sensitive; reattach only slowly to hard substrate in aquarium even if not injured. In the aquarium, specimens soon hide column between or under stones or in holes in substrate (such as dead barnacles). Addition of MgCl2 to aquarium water (for relaxation) provoked spawning in one specimen collected in February 1998 at Quellón.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |