
Lycenchelys tohokuensis Anderson & Imamura, 2002
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4762.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:BEBD8F0D-1347-4A44-86D4-2915433D2E7B |
|
DOI |
https://doi.org/10.5281/zenodo.3809757 |
|
persistent identifier |
https://treatment.plazi.org/id/006C5E1A-FFA0-FFAA-3EC6-B599FEFFADC2 |
|
treatment provided by |
Plazi |
|
scientific name |
Lycenchelys tohokuensis Anderson & Imamura, 2002 |
| status |
|
Lycenchelys tohokuensis Anderson & Imamura, 2002 View in CoL
(Japanese name: Kitagawa-hebigenge)
( Figs. 44–49 View FIGURE 44 View FIGURE 45 View FIGURE 46 View FIGURE 47 View FIGURE 48 View FIGURE 49 ; Table 11)
Lycenchelys tohokuensis Anderson & Imamura, 2002: 355 View in CoL , figs. 1–2 (original description, type locality: off Fukushima Prefecture, Pacific coast of Honshu Island, Japan); Anderson & Fedorov, 2004: 20 (species list); Shinohara & Anderson, 2007: 63 (key to species); Kitagawa et al., 2008: 95, unnumbered fig. (brief description); Shinohara et al., 2009: 724 (species list); Amaoka et al., 2011: 317, unnumbered fig. (brief description); Balushkin et al., 2011: 1026 (species list); Hatooka, 2013: 1227, 2078, unnumbered fig. (key to species); Yamanaka & Ito, 2014: 4 (species list); Nakabo & Hirashima, 2015: 217 (species list and etymology of scientific name).
Materials examined
Holotype: HUMZ 156757 , male, 269.8 mm SL, off Fukushima Prefecture, Tohoku District, northwestern Pacific (37°31.55’N, 142°12.69’E to 37°32.94’N, 142°13.31’E), 697–705 m depth, 18 Apr. 1998, T/V Tanshu-maru, otter trawl. GoogleMaps
Paratypes (4 specimens, 195.3–265.1 mm SL, all from Tohoku District, northwestern Pacific): HUMZ 152369, 1 female, 259.8 mm SL, off Aomori Prefecture (40°42.42’N, 142°09.06’E to 40°41.04’N, 142°09.95’E), 661–667 m depth, 22 Oct. 1997, R/V Wakataka-maru, otter trawl; HUMZ 171668, 1 male, 195.3 mm SL, off Fukushima Prefecture, (36°42.43’N, 141°26.10’E to 36°47.68’N, 141°27.22’E), 582–591 m depth, 11 June 2000, R/V Wakatakamaru, otter trawl; NSMT-P 64042, 1 female, 237.9 mm SL, off Fukushima Prefecture (36°36.25’N, 141°18.23’E to 36°38.91’N, 141°23.67’E), 557–599 m depth, 11 June 2000, R/V Wakataka-maru, otter trawl; HUMZ 178280, 1 female, 265.1 mm SL, off Fukushima Prefecture (37°00.94’N, 141°40.59’ E to 36°57.83’N, 141°36.70’ E), 543–546 m depth, 19 Oct. 2001, R/V Wakataka-maru, otter trawl.
Other specimens (3 specimens): HUMZ 177071, 192808, 206820, 1 male and 2 females, 208.1–270.1 mm SL, Tohoku District, northwestern Pacific.
Diagnosis. Vertebrae 27–29 + 88–91 = 116–119; head length 13.4–14.4% SL; interorbital pore 1; occipital pores 3; postorbital pores 5; suborbital pores 5 + 2; preoperculomandibular pores 8; vomerine teeth 7–11; palatine teeth 8–15, arranged in 2 rows anteriorly and single row posteriorly; opercular flap well developed; pelvic-fins absent; two complete lateral lines, positioned mediolaterally and ventrally; scales present on pectoral fin and its base; body uniformly brown when fresh.
Description. Counts and proportional measurements in Table 11.
Body elongate, cross section oval anteriorly, compressed laterally near tail; its width at anal-fin origin 4.2–5.6 (5.9)% SL. Head relatively short, ovoid; dorsal profile of head gently sloping to above upper edge of gill opening or dorsal-fin origin (above upper edge of gill opening). Snout short, 99.0–142.4 (99.0)% of eye diameter. Eye rounded or slightly oval (rounded), relatively small. Interorbital space moderately narrow, width 23.8–45.4 (23.8)% of eye diameter. Nostril tube long, reaching upper lip when depressed. Mouth subterminal. Posterior edge of upper jaw reaching to about vertical through posterior margin of pupil. Labial lobe of lower jaw developed. Teeth on jaws, vomer and palatine small and conical; upper jaw with 2–3 rows anteriorly and single row posteriorly; lower jaw with 2–3 or 3–4 irregular rows (3–4) anteriorly and single row posteriorly; vomerine teeth irregularly arranged; palatine teeth in 2 rows anteriorly and single row posteriorly. Lower edge of gill opening reaching to or slightly above lower end of pectoral-fin base (above lower end of pectoral-fin base). Opercular flap well developed. Gill rakers, short; those on upper limb triangular, many blunt and some triangular rakers on lower limb ( Fig. 45 View FIGURE 45 ). Pseudobranch filaments relatively long. Two lateral lines, deciduous, complete; mediolateral line origin above about middle portion of abdomen and terminating on tail; ventral lateral line origin posterior to last postorbital pore and terminating on tail. Scales small and cycloid, present on body, pectoral axilla, about basal quarter of pectoral fin, pectoral-fin base, tail and most regions of vertical fins except near margins. Nape with few scales or naked (with few scales). Head without scales.
Dorsal-fin origin nearly above pectoral-fin base; 1st dorsal-fin pterygiophore between neural spines of 3rd and 4th vertebrae. Anal-fin origin below 23rd to 25th (24th) dorsal-fin ray; 1st anal-fin pterygiophore posterior to parapophysis of ultimate or penultimate (penultimate) abdominal vertebra. Last dorsal-fin pterygiophore between neural spines of 3rd to 5th (between 4th and 5th) preural vertebrae. Last anal-fin pterygiophore between hemal spines of 2nd to 4th (between 2nd and 3rd) preural vertebrae. Caudal fin with 2 epural, 4 upper hypural and 4 lower hypural rays. Pectoral fin moderately short, reaching to anterior or middle portion of abdomen; its posterior margin rounded dorsally and notched ventrally. Upper end of pectoral-fin base slightly below lateral midline of body. Pelvic fins absent.
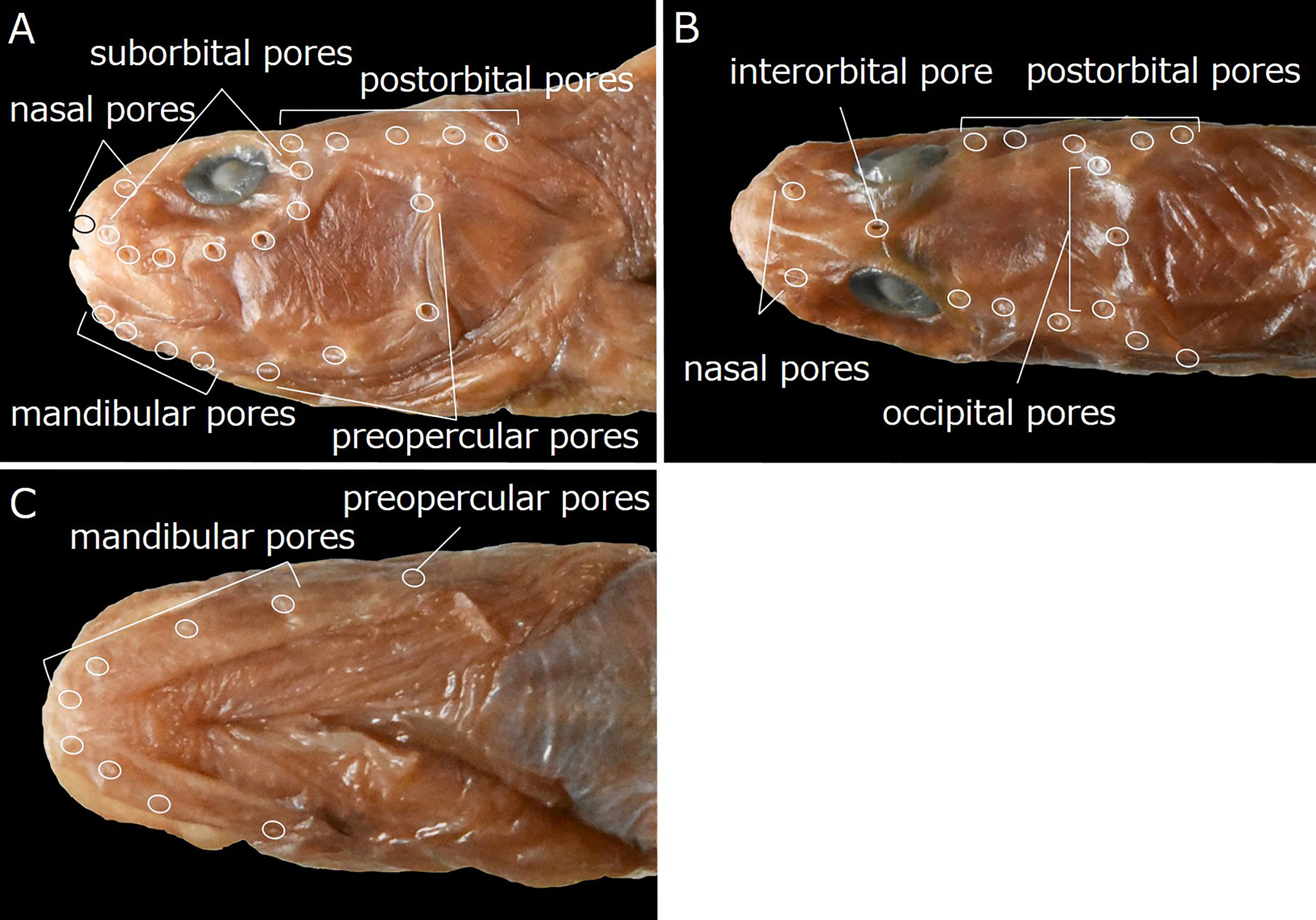
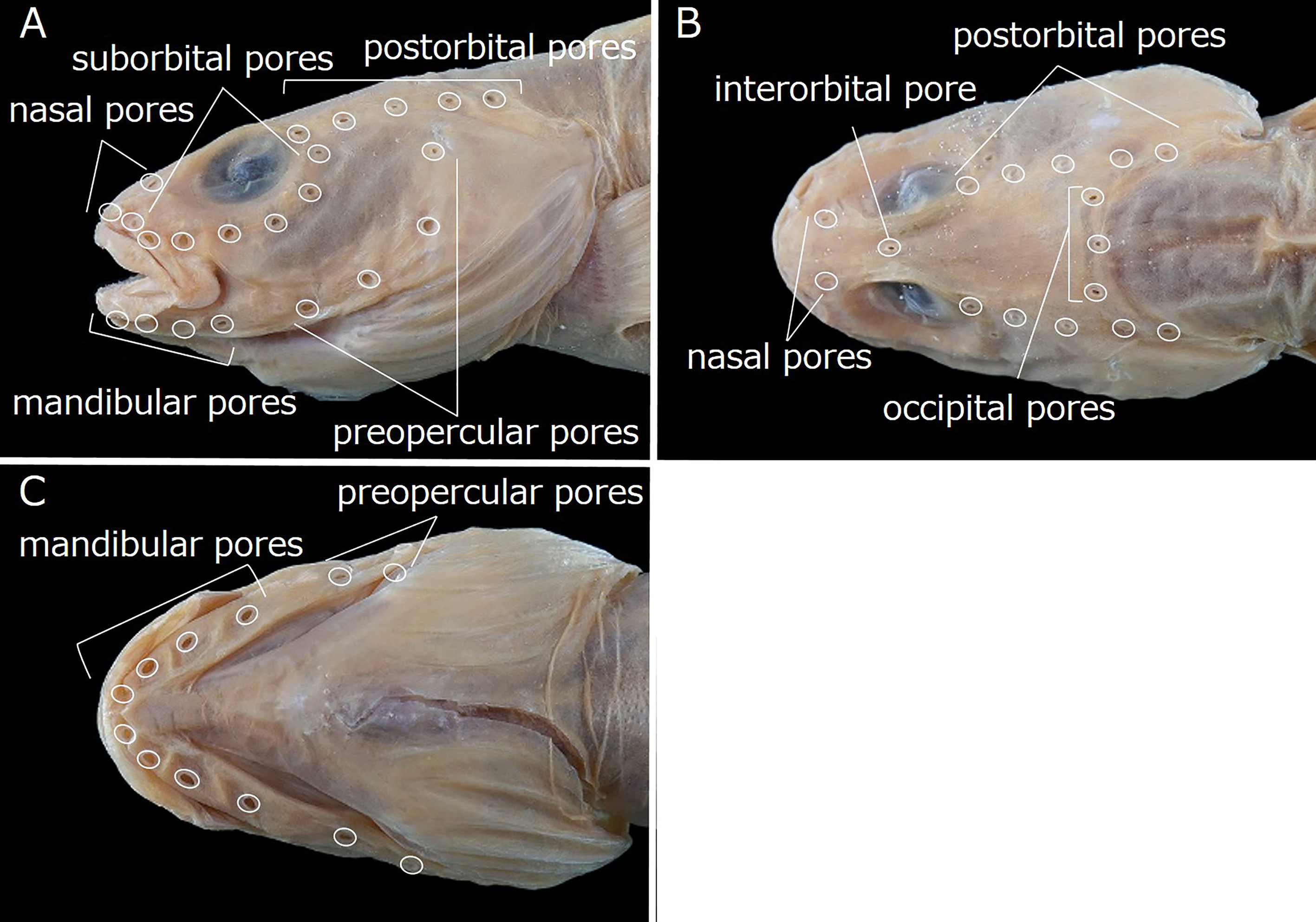
Head pores well developed and distinct. Nasal pores 2; anterior pore in front of nostril tube, posterior pore above 1st suborbital pore ( Fig. 46A, B View FIGURE 46 ). Postorbital pores 5 ( Fig. 46A, B View FIGURE 46 ); holotype with extremely small additional pore posterior to 5th pore on right side. Suborbital pores 7; 5 pores below eye and 2 pores on ascending part of suborbital canal behind eye; 5th pore behind vertical through posterior margin of eye ( Fig. 46A View FIGURE 46 ). Preoperculomandibular pores 8; 4 on lower jaw and 4 on preopercle; last preoperculomandibular pore posterior to lower margin of eye ( Fig. 46A, C View FIGURE 46 ). One interorbital pore on dorsal midline between anterior margins of pupils (anterior margin of pupils) ( Fig. 46B View FIGURE 46 ). Occipital pores 3; 1 on dorsal midline of occiput, and remaining 2 on either side; 3 pores transversely aligned or middle pore slightly posterior to those on either side (middle pore located slightly behind side pores); all pores located anterior to 4th postorbital pore ( Fig. 46B View FIGURE 46 ).
Color in alcohol. Holotype ( Fig. 47 View FIGURE 47 ) with brownish head and uniformly paler body, pectoral fin and vertical fins; dark brown margins of pectoral and vertical fins and grayish abdomen. Body purplish brown in HUMZ 171668. Coloration of other specimens similar to holotype.
Color when fresh (based on color photograph of HUMZ 152369; Fig. 44 View FIGURE 44 ). Head, body and vertical fins uniformly brown, pectoral fin paler; margins of pectoral and vertical fins, and opercular region blackish; abdomen grayish.
Distribution. Off northwestern Pacific coast of Tohoku District from Aomori to Fukushima prefectures, at depths of 543–905 m ( Anderson & Imamura, 2002; Anderson & Fedorov, 2004; Shinohara & Anderson, 2007; Kitagawa et al., 2008; Shinohara et al., 2009; Amaoka et al., 2011; Hatooka, 2013; Yamanaka & Ito, 2014; this study).
Size. The largest specimen examined during this study measured 270.1 mm SL (HUMZ 192808, 275.2 mm TL), about equal to the previously recorded maximum length (HUMZ 156757, holotype, 270 mm SL) ( Anderson & Imamura, 2002).
Remarks. Other than L. tohokuensis , only five species of Lycenchelys are known to lack pelvic fins: Lycenchelys fedorovi Anderson & Balanov, 2000 , Lycenchelys hureaui ( Andriashev, 1979) , Lycenchelys monstrosa Anderson, 1982 , Lycenchelys nanospinata Anderson, 1988 and Lycenchelys xanthoptera Anderson, 1991 ( Anderson, 1982a, 1988, 1991; Anderson & Balanov, 2000; Anderson & Imamura, 2002). Lycenchelys tohokuensis is distinguished from the five species in having two lateral lines and palatine teeth arranged in two rows anteriorly (vs. having a single lateral line in all five species and palatine teeth arranged in a single row in L. fedorovi , L. hureaui , L. monstrosa and L. nanospinata , or lacking palatine teeth in L. xanthoptera ) ( Anderson, 1982a, 1988, 1991; Anderson & Balanov, 2000; Anderson & Imamura, 2002). Lycenchelys tohokuensis also differs from L. fedorovi , L. hureaui , L. monstrosa and L. nanospinata in having 116–119 total vertebrae (vs. 132–139 in L. fedorovi , 104–112 in L. hureaui , 126–132 in L. monstrosa and 105 in L. nanospinata ), from L. hureaui , L. monstrosa , L. nanospinata and L. xanthoptera in having 1 interorbital pore and 3 occipital pores (vs. interorbital pores and occipital pores absent in the four), and from L. monstrosa in having vomerine teeth (vs. usually absent in L. monstrosa ) ( Anderson, 1982a, 1988, 1991; Anderson & Balanov, 2000; Anderson & Imamura, 2002; this study).
Although the Lycodinae are recognized as having an L-shaped series of suborbital bones ( Anderson, 1994), they are arranged in a semicircular series in L. tohokuensis ( Fig. 45A View FIGURE 45 ). The latter configuration also occurs in the other three subfamilies: Lycozoarcinae Andriashev, 1939 , Zoarcinae Gill, 1862 and Gymnelinae Gill, 1864 ( Anderson, 1994) . Lycenchelys tohokuensis clearly differs from species of Lycozoarcinae (including only Lycozoarces regani Popov, 1933 ) and Zoacinae (only Zoarces Cuvier, 1829 with six species) in having scales, and teeth on the vomer and palatine, and lacking the pelvic fins and dorsal-fin spines (vs. having pelvic fins and lacking scales in the Lycozoarcinae , and having pelvic fins and dorsal-fin spines, and lacking vomerine and palatine teeth in the Zoarcinae ) ( Anderson, 1994). When compared with genera of Gymnelinae , L. tohokuensis mostly resembles Ericandersonia Shinohara & Sakurai, 2006 [only 1 species, Ericandersonia sagamia Shinohara & Sakurai, 2006 ( Fig 48 View FIGURE 48 )] in having similar numbers of dorsal-fin rays (112–116 vs. 113–116), anal-fin rays (93–94 vs. 93–95), pectoral-fin rays (14 vs. 14), and total vertebrae (116–119 vs.117–118), respectively ( Anderson & Imamura, 2002; Shinohara & Sakurai, 2006; this study). Furthermore, numbers and arrangements of head pores in the two species are the same (2 nasal, 5 postorbital, 7 suborbital, 4 mandibular, 4 preopercular, 1 interorbital and 3 occipital pores) ( Figs. 46A, B, C View FIGURE 46 , 49A, B, C View FIGURE 49 ) ( Anderson & Imamura, 2002; Shinohara & Sakurai, 2006; this study). The two are distinguishable, however, by the presence of 2 lateral lines, and a developed labial lobe on the lower jaw, and the lower edge of the gill opening set slightly above the lower end of the pectoral-fin base in L. tohokuensis (vs. lateral line single and mediolateral, labial lobe of lower jaw absent, and lower edge of the gill opening not reaching lower end of pectoral-fin base in E. sagamia ) ( Anderson & Imamura, 2002; Shinohara & Sakurai, 2006; this study). In addition, E. sagamia is characterized by the presence of a weak ridge (= pseudosubmental crest sensu Shinohara & Sakurai, 2006) on the ventral surface of the mandibula, which is absent in other zoarcid species ( Shinohara & Sakurai, 2006; this study).
The question of retaining Lycenchelys tohokuensis in Lycenchelys because of its peculiar arrangement of the suborbital bones, relative to that of other members of the subfamily needs to be tested by a phylogenetic study of the entire group.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lycenchelys tohokuensis Anderson & Imamura, 2002
| Kawarada, Shumpei, Imamura, Hisashi, Narimatsu, Yoji & Shinohara, Gento 2020 |
Lycenchelys tohokuensis
| Nakabo, T. & Hirashima, Y. 2015: 217 |
| Yamanaka, T. & Ito, K. 2014: 4 |
| Hatooka, K. 2013: 1227 |
| Amaoka, K. & Nakaya, K. & Yabe, M. 2011: 317 |
| Balushkin, A. V. & Sheiko, B. A. & Fedorov, V. V. 2011: 1026 |
| Shinohara, G. & Narimatsu, Y. & Hattori, T. & Ito, M. & Takata, Y. & Matsuura, K. 2009: 724 |
| Kitagawa, D. & Imamura, H. & Goto, T. & Ishito, Y. & Fujiwara, K. & Ueda, Y. 2008: 95 |
| Shinohara, G. & Anderson, M. E. 2007: 63 |
| Anderson, M. E. & Fedorov, V. V. 2004: 20 |
| Anderson, M. E. & Imamura, H. 2002: 355 |