
SAEUNGAE, Taai & Harbach, 2015
|
publication ID |
https://doi.org/ 10.1111/zoj.12236 |
|
persistent identifier |
https://treatment.plazi.org/id/C46F9003-FFAD-FFE6-C30C-ACBEFCCF1533 |
|
treatment provided by |
Carolina |
|
scientific name |
SAEUNGAE |
| status |
sp. nov. |
ANOPHELES View in CoL ( ANOPHELES ) SAEUNGAE SP. NOV.
Anopheles (Anopheles) barbirostris View in CoL in part of Reid, 1962 ( Thailand, ♀ * ♂ E* L P*, taxonomy); Harrison & Scanlon, 1975 ( Thailand, ♀ * ♂ * L* P*, taxonomy); Reid et al., 1979 ( Thailand, ♀ L P morphology).
Anopheles barbirostris View in CoL in part (?) of Harrison et al., 1988 ( Thailand, A L P morphology).
Anopheles barbirostris forms A (in part) and B (in part) of Baimai et al., 1995 ( Thailand, metaphase karyotype).
Anopheles barbirostris Forms A and B (in part) of Saeung et al., 2007 ( Thailand, mitotic karyotype, crossmatings, COI and COII mtDNA, ITS2 rDNA).
Anopheles barbirostris View in CoL species A2 of Saeung et al., 2008 ( Thailand, mitotic karyotype, cross-matings, COI and COII mtDNA, ITS2 rDNA); Suwannamit et al., 2009 ( Thailand, cross-matings, metaphase karyotype, COI and COII mtDNA, ITS2 rDNA); Thongsahuan et al., 2011 ( Thailand, Plasmodium susceptibility); Otsuka, 2011 ( Thailand, ITS2 rDNA).
Anopheles barbirostris View in CoL Clade IV (unknown species) of Paredes-Esquivel et al., 2009 (Sumatran Indonesia, Thailand, COI mtDNA, ITS2 rDNA); Paredes-Esquivel & Townson, 2014 (Sumatran Indonesia, Thailand, ITS2 rDNA).
Diagnosis
Adult females of An. saeungae resemble the adult females of An. campestris in having a similar pattern of pale scales on the abdominal sterna, but the scaling of the wings is paler, as in An. barbirostris . This combination of characters, along with COI and ITS2 sequences (see below), readily distinguish An. saeungae from other species of the Barbirostris Complex. Some additional potentially differential morphological characters are denoted below, but for practical purposes An. dissidens is nearly isomorphic with other species of the complex.
Description
Female
Overall similar to An. barbirostris , but generally smaller, as indicated by measurements of the proboscis, maxillary palpus, antennal flagellum, wing, and forefemur listed in Table 1: lengths of proboscis, maxillary palpus, wing, vein R 4+5, forefemur, and width of wing statistically significantly shorter ( Table S2); apical pale band of hindtarsomere 3 0.04–0.18 mm (mean 0.10 mm), extended across joint onto base of hindtarsomere 4 in 20% of females examined.
Male
As described for An. barbirostris but generally smaller, as indicated by measurements of the gonocoxite, distal parabasal setae, internal seta, gonostylus, claspette club, and aedeagus listed in Table 2, but means of measurements not significantly different ( Table S3).
Pupa
As described for An. barbirostris , indistinguishable; setal branching in Table S8; branching of seta 2 compared with other species of the complex in Table 3.
Larva, fourth instar
As described for An. barbirostris ; setal branching in Table S9; differing in having seta 13-II with eight to 13 (eight) branches; sum of branches of pair of 13- P = 11–18, pair of seta 10-III = 4, 5 (4).
Mitotic karyotype
Three types of X chromosome (X 1, X 2, X 3) and two types of Y chromosome (Y 1, Y 2) comprising two karyotypic forms (X 1 X 2 X 3 Y 1, X 1 X 2 X 3 Y 2) have been identified in the early fourth-instar larval brains of An. saeungae ( Baimai et al., 1995; Saeung et al., 2008; Suwannamit et al., 2009).
Cross-matings
The results of crosses between An. saeungae (as An. barbirostris species A2) and An. dissidens (as species A1) are summarized under An. dissidens above. Suwannamit et al. (2009) obtained similar results from cross-matings between An. saeungae (as species A2) and An. barbirostris (as species A4), i.e. incomplete hatching of eggs, asynaptic polytene chromosomes in larvae, reduced pupation, and abnormal reproductive organs in a few adults.
DNA sequence
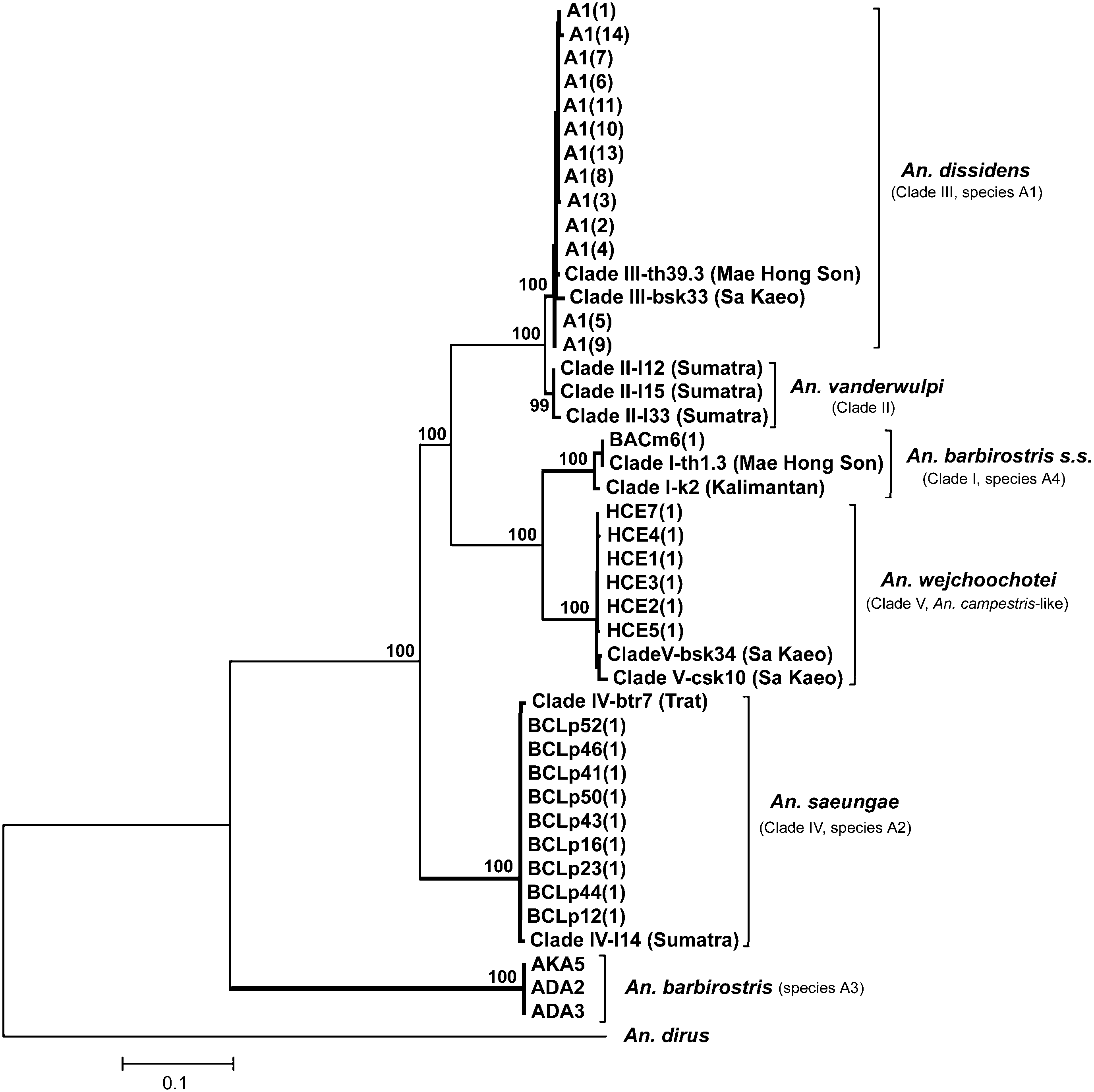
Specimens identified as An. saeungae are shown in Table S1, together with GenBank accession numbers for ITS2 and COI sequences. The ITS2 subunit for An. saeungae yields a dominant product of 1678 bp. The six interspecifically variable sites of the COI gene at bases 31, 82, 91, 181, 433, and 631 that are unique for this species are shown in Figure 4 View Figure 4 . The results of Bayesian analyses of ITS2 and COI sequences are shown in Figures 5 View Figure 5 and 6 View Figure 6 , respectively. Both trees show that An. saeungae is a distinct member of the Barbirostris Complex. Our ITS2 sequences for An. saeungae (BCLp) fall within a strongly supported clade ( Fig. 5 View Figure 5 , BPP 100%) with two sequences (Btr7 and l14) of An. barbirostris Clade IV of Paredes-Esquivel et al. (2009).
Bionomics
Adult females of An. saeungae have been collected in buffalo-baited traps, but are likely to attack humans on occasions when preferred hosts are scarce. Thongsahuan et al. (2011) found that 6.67% of females (as An. barbirostris species A2) had developed sporozoites of P. vivax during experimental infection studies.
Distribution
Based on COI, COII, and ITS2 sequences, An. saeungae is currently only definitely known to occur in Indonesia (Sumatra) and Thailand (Chanthaburi, Lampang, Phetchaburi, Ratchaburi, Sa Kaeo, Trat, Ubon Ratchaburi, and Udon Thani Provinces) ( Saeung et al., 2008; Paredes-Esquivel et al., 2009; Suwannamit et al., 2009; present study).
Etymology
This species is named in honour of Dr Atiporn Saeung ( Department of Parasitology , Faculty of Medicine, Chiang Mai University, Chiang Mai, Thailand) for her many contributions to our knowledge of mosquitoes in Southeast Asia, especially her cytogenetic and molecular studies of the Barbirostris Group, which provided information for further studies of these economically important mosquitoes .
Type series
Two-hundred and ninety-seven specimens (135 ♀, 67 ♂, 36 Le, 35 Pe, 24 L) derived from nine molecularly identified progeny broods: BCLp12(1), BCLp16(1), BCLp23(1), BCLp41(1), BCLp43(1), BCLp44(1), BCLp46(1), BCLp50(1), and BCLp52(1). Holotype, ♀ [BCLp16(1)-2], with Le and Pe on microscope slide, offspring of female collected as follows: THAILAND, Lampang Province, Ko Kha District , Ban Don Tham , buffalo-baited trap, 7.xii.2013, coll. Choochote et al. Paratypes, same data as holotype : 20 ♀ LePe [BCLp12(1)-3 to -6, -8, -10, -12; BCLp16(1)-1, -5 to -8, -12; BCLp23(1)-1, -3, -4, -8 to -11] ; 1 ♀ Le [BCLp12(1)- 7] ; 113 ♀ [BCLp12(1)-14, -16, -17, -21, -23, -25 to -29, -32; BCLp16(1)-13, -17, -18, -20, -22 to -49; BCLp23(1)- 13, to -15, -17 -19; BCLp41(1)-1 to -3, -7, -8, -10, -11; BCLp43(1)-8 to -11; BCLp44(1)-2, -3, -5, -7, -9, -11 to -23, -100; BCLp46(1)-2, -3, -5, -7, -12, -14, -15; BCLp50(1)-1, -3, -9 to -11, -13, -14, -16 to -22; BCLp52(1)-6 to -19]; 14 ♂ LePe [BCLp12(1)-1, -2, -9, -11; BCLp16(1)-3, -4, -9 to -11; BCLp23(1)-2, -5 to -7, -12] ; 53 ♂ [BCLp12(1)-13, -15, -18 to -20, -22, -24, -30, -31; BCLp16(1)-14 to -16, -19, -21; BCLp23(1)-16, -18, -20 to -22; BCLp41(1)-4 to -6, -9; BCLp43(1)-1 to -7; BCLp44(1)-1, -4, -6, -8, -10; BCLp46(1)-1, -4, -6, -13, -16; BCLp50(1)-2, -4 to -8, -12, -15; BCLp52(1)-1 to -5]; 24 L [BCLp12(1)-A, -B; BCLp16(1)-A, -B; BCLp23(1)- A, -B; BCLp41(1)-A, -B; BCLp43(1)-A, -B; BCLp46(1)- A, -B; BCLp50(1)-A, -B, -C, -D, -E, -F; BCLp52(1)-A, -B,-C, -D, -E, -F]. The type series is deposited in BMNH .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.