
Vespina meridiana Hirowatari & Yagi, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4927.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:96B9981B-01B5-4828-A4C6-E2E4A08DB8F2 |
|
DOI |
https://doi.org/10.5281/zenodo.4565599 |
|
persistent identifier |
https://treatment.plazi.org/id/E83387BF-FFA0-FF8B-5094-B539FE751E9E |
|
treatment provided by |
Plazi |
|
scientific name |
Vespina meridiana Hirowatari & Yagi |
| status |
sp. nov. |
Vespina meridiana Hirowatari & Yagi , sp. nov.
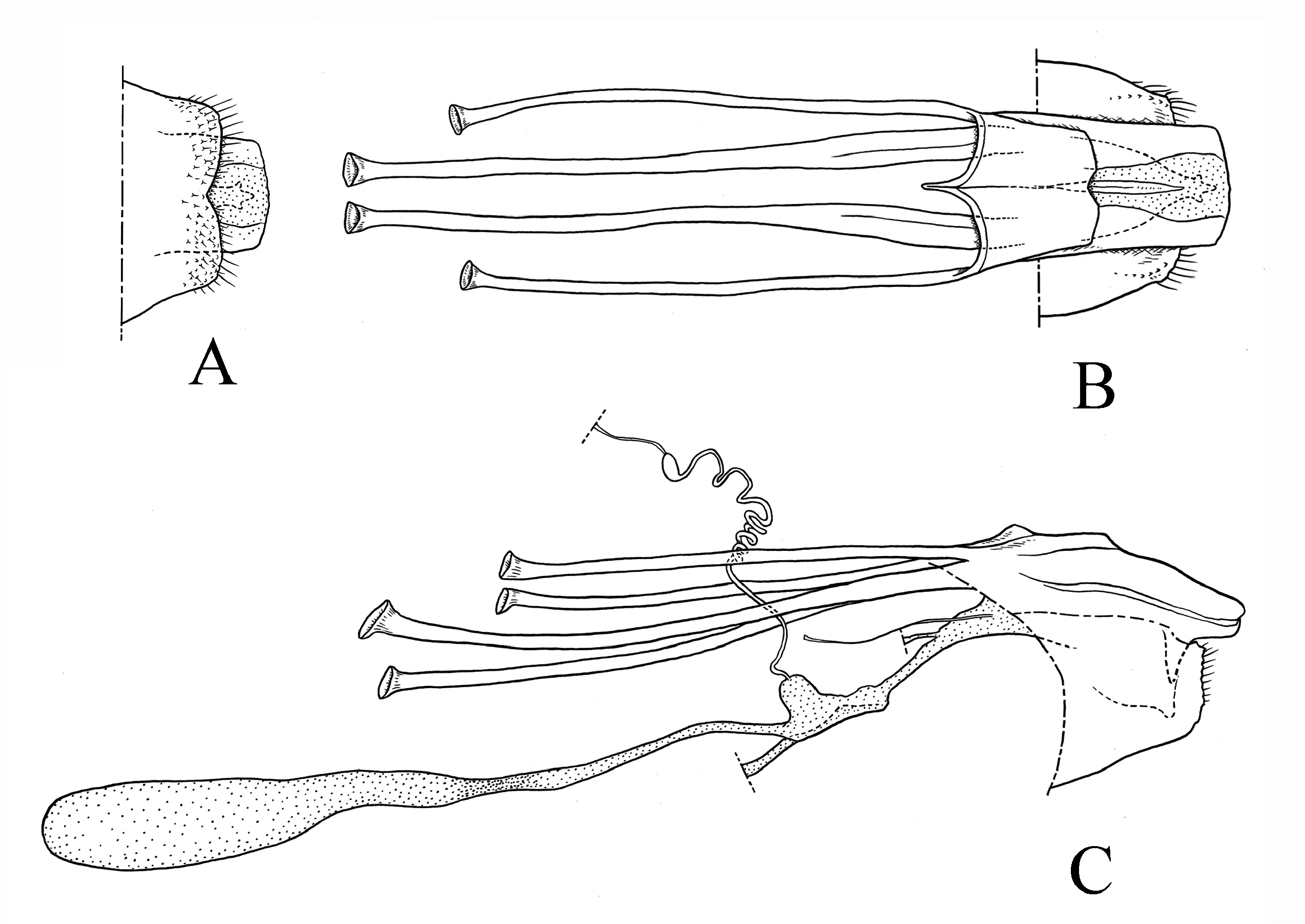
( Figs 1D View FIGURE 1 , 2D View FIGURE 2 , 3D View FIGURE 3 , 4–6 View FIGURE 4 View FIGURE 5 View FIGURE 6 , 13C, D View FIGURE 13 )
Type material: holotype ♀, “ Japan [Ryukyu]/ Yonahadake / Kunigami-son / Okinawa Pref. / 21.iv.2004 / T. Ueda ”, “ Host : / em. 28.VI. 2004 ” Sample ID SaY 89 in OPU .
Other material. Larvae: 6 exs 4–6.iv.1996, Hatsuno, Amami-Oshima, Kagoshima Pref.; 10 exs 22–24.iii.2002, Kunigami-son, Okinawa Pref., T. Hirowatari ( OPU); 20 exs 15–18.iii.2017, Hentona–Okuni-rindo, T. Hirowatari & S. Yagi, Sample ID SaY90, 91 ( ELKU); 2 exs Benoki, 19.iii.2020, S. Yagi, DNA extraction number No. SaY448 ( ELKU) (some of them were preserved in ca. 80 or 100% ethanol; all collected on Morella rubra , Myricaceae ).
Diagnosis. Difficult to distinguish from other Asian Vespina species externally, but may be distinguishable by the entirely pale yellow hairs of the head ( Fig. 2D View FIGURE 2 ); in V. nielseni , the yellow hairs are mixed with orange hairs on the vertex ( Fig. 2A, B View FIGURE 2 ), whereas in V. sichuana , they are mixed with light-brown hairs on the posterior side of the head ( Fig. 2E, F View FIGURE 2 ). The triangular creamy-white tornal spot on the forewing is absent (well-developed in V. nielseni and indistinct in V. sichuana ). In the female genitalia, the anterior end of the apophysis posterioris is rather small, 2–2.5 × as wide as the base, and the ductus bursae is relatively long, as long as the corpus bursae.
Description.
Male unknown.
Female ( Figs 1D View FIGURE 1 , 2D View FIGURE 2 , 3D View FIGURE 3 ). Wingspan 10.0 mm in holotype. Forewing length 4.7 mm in holotype.
Head: Vertex and frons with pale yellow hairs, entirely rough; antenna 2/3 the length of the forewing. Ocellus absent. Compound eye large. Maxillary palpus short (slightly shorter than the labial palpus), pale yellow mixed with pale brown. Labial palpus porrect, short and slender, pale yellow, second segment with several distal brown bristles.
Thorax: Tegula, prothorax and mesothorax dorsally dark brown with a weak purple gloss. Metathorax with dark brown scales on the posterior. Foreleg dark brown, mixed with pale yellow on the femur; midleg dark brown, mixed with pale yellow on the femur, with blackish brown tibial spurs; hindleg dorsally dark brown and partly mixed with pale yellow, with pale yellow hairs ventrally on the tibia, and pale yellow tibial spurs. Wings narrow. Forewing brown. Hindwing brown. Frenulum absent or reduced with approximately 15 thin pseudofrenular bristles at the costal margin.
Abdomen: Brown with golden luster.
Female genitalia ( Fig. 4 View FIGURE 4 ): Apophyses stout, each anterior end swollen, trumpet-shaped; apophysis posterioris slightly longer than the apophysis anterioris, with the anterior end rather small and the posterior half depressed and expanded laterally. Apex of ovipositor dorsoventrally flattened with one median semicircular projection. Spermathecal vesicle small and oval. Corpus bursae elongate, ellipsoidal; signa absent. Ductus bursae relatively long, as long as corpus bursae, weakly sclerotized near the caudal portion of the corpus bursae.
Immature stages ( Figs 5 View FIGURE 5 , 6 View FIGURE 6 ).
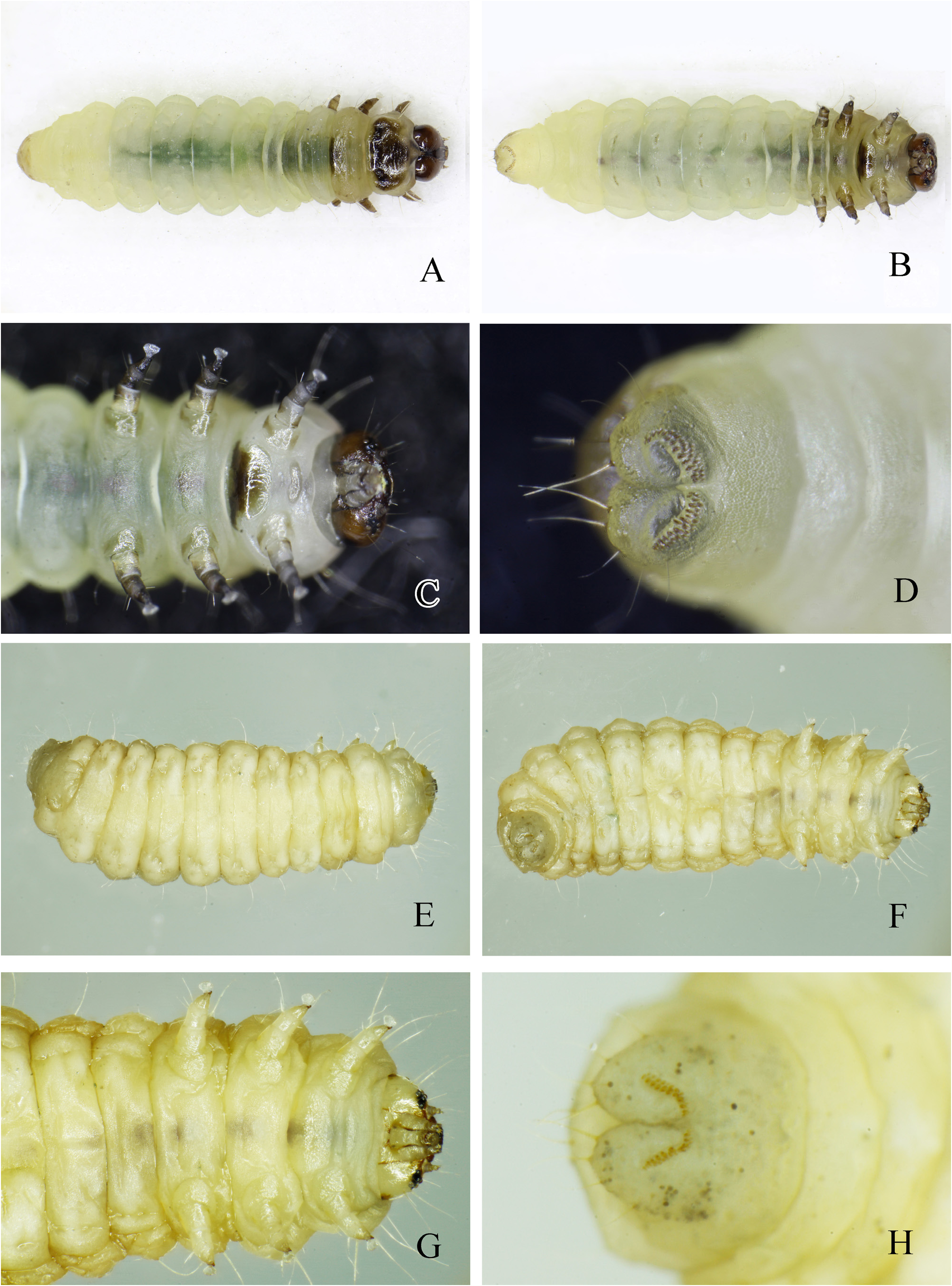
Pre-mature larva ( Figs 5H View FIGURE 5 , 6 View FIGURE 6 A–D). Color: Yellowish-white.
Length: 5.2 –5.7 mm (n = 5). Head: Prognathous with six pairs of stemmata. Head capsule dark brown. Antennae with three sclerotized segments. Thorax: Prothoracic tergite well sclerotized; mesothoracic tergite weakly sclerotized, apart from the well-sclerotized dorsal part of the posterior margin; metathoracic tergite not sclerotized. Prothoracic spiracle distinct, with a sclerotized rim. Thoracic legs well developed; pretarsus with a squamiform seta adjacent to the claw ( Fig. 6C View FIGURE 6 ). Abdomen: No distinctive pigmentation except a weakly sclerotized dorsal plate on the tenth segment. Prolegs reduced; crochets arranged as a transverse band consisting of two rows of spines: 9–12 larger posterior spines associated with 7–9 smaller anterior spines on segments 3, 4, 5, and 6, alongside two or three rows of 10–15 transverse spines on segment 10 ( Fig. 6D View FIGURE 6 ). Number of crochets variable. Spiracles oval on the first to eighth segments.
Mature larva ( Fig. 6 View FIGURE 6 E–H). Color: Yellowish-white.
Length: 4.2–4.4 mm (n = 3).
Body: Stout and fat. Head: Head capsule pale brown with brown area around the base of the antennae and the six pairs of stemmata; enclosed area of stemmata blackish-brown. Head capsule flattened. Posterior margin more bluntly V-shaped than that of pre-mature larva. Antenna with three weakly sclerotized segments. Thorax: Thoracic tergites not well-sclerotized. Prothoracic spiracle not pigmented. Pretarsus with a squamiform seta adjacent to the claw, almost the same size as that of the pre-mature larva. Abdomen: No distinctive pigmentation on the yellowish-white abdomen except a very weakly sclerotized dorsal plate on the tenth segment. Prolegs reduced; crochets with a transverse band consisting of a row of 7–9 spines on segments 3, 4, 5, and 6, and a row of 12–15 transverse spines on segment 10 ( Fig. 6H View FIGURE 6 ) crochets evidently less than that of the pre-mature larva. Spiracles same as in the pre-mature larva.
Pupa: Not examined.
Etymology. The specific name is derived from the Latin meridiana (=southern), in reference to the southernmost distribution of the new species in the genus.
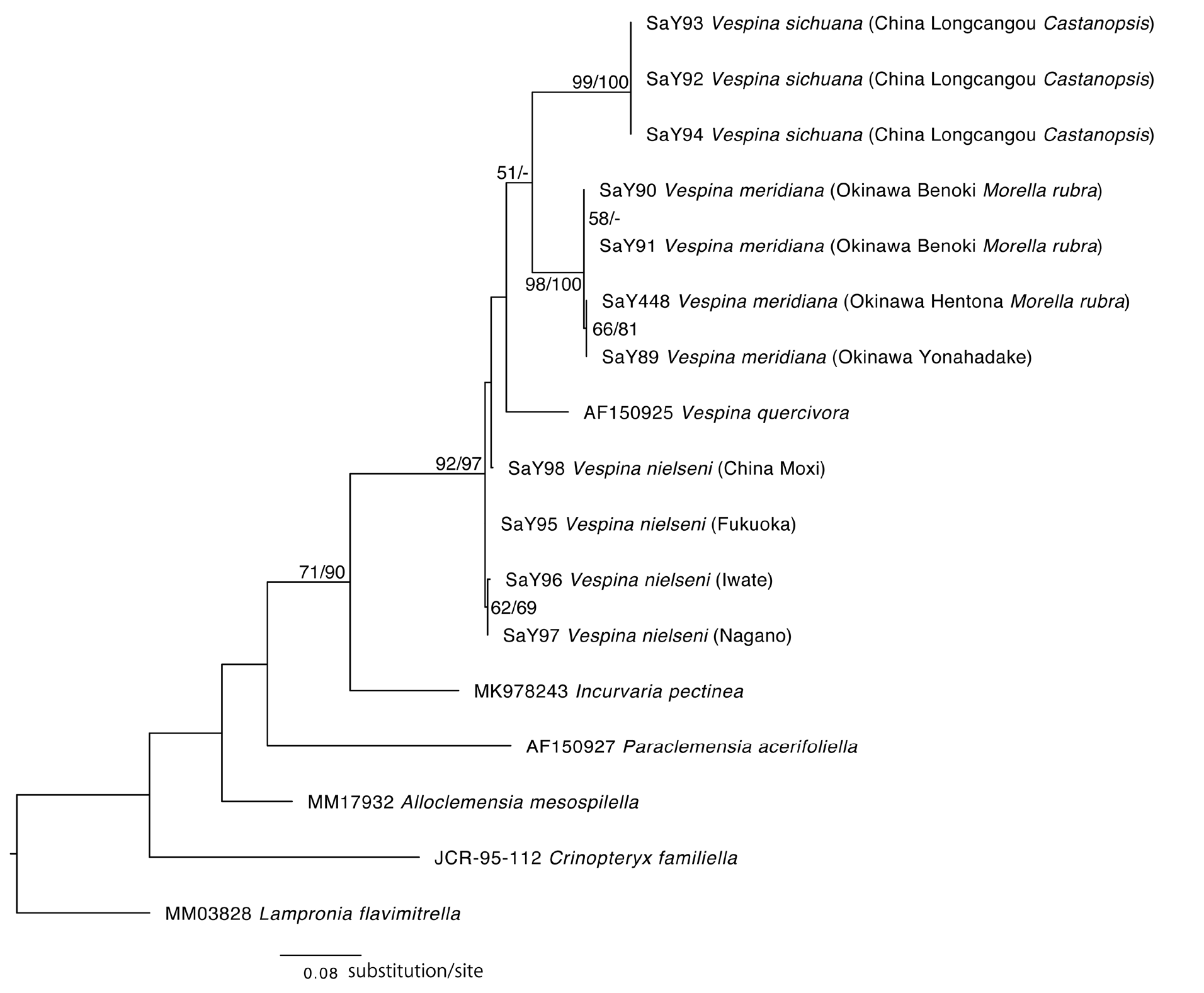
Barcode data. DNA barcodes of four specimens were generated ( Table 1). A single female specimen (SaY89; Fig. 14 View FIGURE 14 ) collected at Yonahadake, Kunigami-son, Okinawa Pref. —the holotype of Vespina meridiana ( Fig. 1D View FIGURE 1 )— showed 0.46–1.86% (SaY90, SaY91, SaY448) pairwise distance (in terms of the sequenced 658 bp COI barcoding region) from three larval specimens collected at Benoki and Hentona, Kunigami-son on Morella rubra leaves ( Table 3). These distances were smaller than typical empirical interspecific distances (e.g., Hebert et al. 2003). Therefore, we concluded that the female from Yonahadake and the larvae from Morella rubura were the same species. The specimens collected from the two larger areas in the northern part of Okinawa Island (Benoki vs. Hentona and Yonahadake, ca. 10 km away) have some large intraspecific distances (1.70–2.02%), while the pairwise distances within the close areas (Yonahadake vs. Hentona) have small intraspecific distances (0.46%). Perhaps this represents the low migratory habits of adults.
Host plant. Myricaceae : Morella rubra .
Biology. In March 2017, we found a young larva of Vespina meridiana shouldering a thin case on a twig of Morella rubra ( Fig. 13D View FIGURE 13 ), perhaps just after constructing the oval case. Later, the larva cut an oval section from the leaf and added it to the top of the old case, in the same manner as other species of Vespina (e.g., V. quercivora and V. nielseni ).
Distribution. Japan (Okinawa Island and Amami-Oshima Island).
Remarks. One of the authors (T. Hirowatari) found portable larval cases of this species on Morella rubra (Myricaceae) on Amami-Oshima Island in March 1996. Since then, we have collected many portable larval cases of this species on the host plant in the northern part of Okinawa (Kunigami-son), and reared many larvae. However, we have failed to obtain pupae and adults under laboratory conditions in Osaka and Fukuoka, Japan. Most of the larvae collected in the spring survived for over four months; however, from spring to summer, many of them died in their pre-mature stage. By the end of summer, a small number of larvae appeared to be in the last instar phase and were alive until the end of the year, at which point they finally succumbed to death.
No adults have been collected in the field. The larva of the holotype female within a portable case on an undetermined plant leaf (probably Morella rubra ) was collected by Dr. Tatsuya Ueda in Okinawa in June 2004. It was in a non-feeding stage before pupation because no trace of feeding remained on the leaf. In the rearing condition, most premature larvae are observed to move from the leaves of the host plant to the wall of the rearing container and fix their case on the wall with a thread. The larva of the holotype also moved in such a manner for pupation. Then, the female emerged one week later. This species seems to undergo diapause before emergence, but the diapauseterminating cue is unclear .





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |