
Valeriana sobraliana Rabuske & Iganci, 2019
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.423.1.2 |
|
DOI |
https://doi.org/10.5281/zenodo.10257665 |
|
persistent identifier |
https://treatment.plazi.org/id/039987E0-FF9E-837B-FF28-FCBEFD4DFD16 |
|
treatment provided by |
Guilherme |
|
scientific name |
Valeriana sobraliana Rabuske & Iganci |
| status |
sp. nov. |
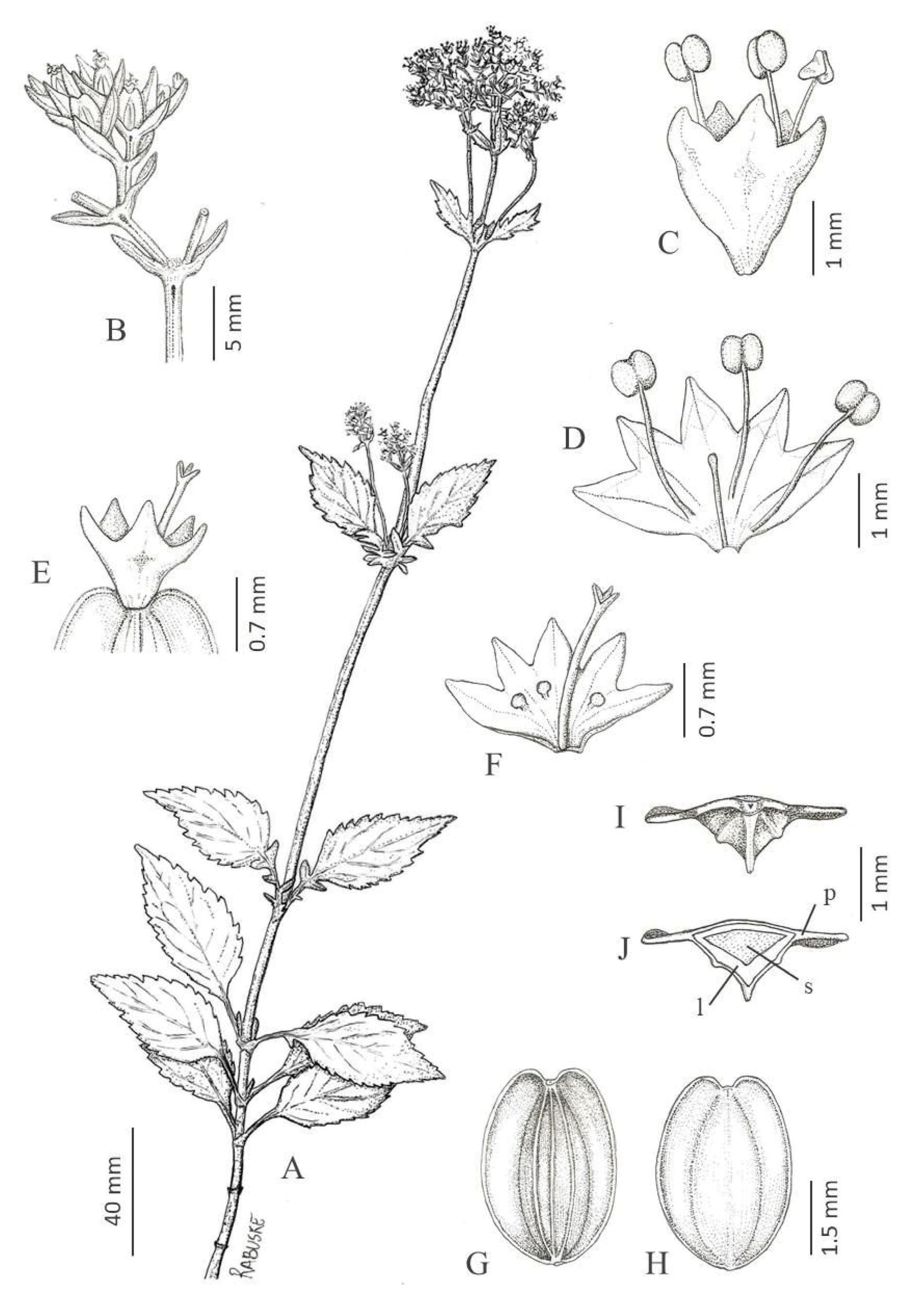
Valeriana sobraliana Rabuske & Iganci View in CoL sp.nov. ( Figures 1 View FIGURE 1 , 2 View FIGURE 2 and 3 View FIGURE 3 )
Type:— Brazil. Santa Catarina: Garuva, Campos do Quiriri , 26°01’56”S 48°58’45”W, 1350 m elev., 31 October 2013, pist. fl., E.Barboza, J.Cordeiro, J.M.Silva & J.T.Motta 4038 (holotype: MBM!, isotypes: FLOR!, FURB!, ICN!, RB!) GoogleMaps .
The new species is morphologically similar to Valeriana catharinensis Graebn. (1899: 427) , V.ulei Graebn. (1899: 436) and V. organensis Gardn. (1845: 112) ( Figure 4 View FIGURE 4 ). It differs mainly by the leaves strongly discolorous, with irregularly crenate-serrate margins and glabrous indument (vs. blades at most slightly discolorous, margins serrate or serrulate, and when irregularly crenate-serrate, always pubescent [ V. catharinensis ]), by the petioles reaching ⅓ to ⅔ the length of the leaf blade (vs. leaves sessile, pseudopetiolate or petiolate; the petioles, when present, never exceeding ⅓ the length of the leaf blade), by the bracteoles lanceolate and larger, 3–3.2 × 1–1.2 mm, with acute to attenuate apex, eventually caudate, base rounded to subcordate (vs. bracteoles ovate or elliptic, 1.5–2.8 × 0.7–1 mm, apex always acute, base rounded or attenuate), by the larger staminate flowers, 3.3 × 2.7 mm, with a pistilodium larger than 1.6 mm long (vs. staminate flowers up to 2.8 × 2.2 mm, the pistilodium up to 1.4 mm long), and by the elliptic achenes (vs. achenes ovate or globose) ( Table 1 View TABLE 1 ).
Dioic subshrub, 30–100 cm tall, perennial, rhizomatous, erect, simple to branched; plants glabrous except for tufts of hyaline trichomes 0.3–0.7 mm long, denser and conspicuous in the nodes or sparse along the axis of the inflorescence. Branches fistulous, terete, 2.5–4.2 mm in diameter, glabrous, striate, corrugated at floral axis; internodes 4–46 mm long at base and 8–115 mm long at floral axis, eventually slightly winged, wings 0.3–0.5 mm wide, inconspicuous, connecting the nodes. Leaves opposite and decussate, sparse along the branches or in distal subrosettes, heteromorphic, the most basal caulinar leaves marcescent and deciduous in late phenophases, simple, blades elliptic to lanceolate, eventually ovate, 17–85 × 8–28 mm, becoming smaller distally, chartaceous or at most slightly coriaceous, strongly discolorous, the adaxial face dark-green, the abaxial face light-green; apex acute to attenuate; base cuneate to attenuate, slightly asymmetrical and falcate; petioles 6–30 × 1–1.5 mm, reaching ⅓ to ⅔ the length of the leaf blade, decurrent; primary vein adaxially sulcate, abaxially salient; secondary veins 3–7 pairs, anastomosing; margin entire, revolute, irregularly crenate-serrate, teeth 5–14 pairs, 1.5–9 × 0.5–3.5 mm, teeth apex apiculate and glandular; apiculum up to 0.2 mm long; teeth glands rounded, concave, 0.2–0.3 mm in diameter; distal leaves eventually pinnatipartite to pinnately lobed; basal lobes 1–3 pairs, 3–11 × 1–3.5 mm, elliptical, elliptic to lanceolate, sometimes hardly visible, becoming larger distally; margin irregularly crenate-serrate, eventually serrate, teeth eventually elongated. Inflorescence terminal in compound dichasial cyme, the distal portion capituliform, with 2–5 pairs of secondary branches (paracladia, sensu Larsen 1986), rachis 20–185 mm long, secondary axes 12–65 mm long, tertiary axes dichasial up to 10 mm long. Bracts on the two lower nodes of the inflorescence, 18–64 × 10–23 mm, similar to upper leaves; distal bracts elliptic to lanceolate, navicular, 6–12 × 2–4 mm. Bracteoles lanceolate, 3–3.2 × 1–1.2 mm, green, acute to attenuate apex, eventually caudate, base rounded to subcordate, decurrent. Flowers of both sexes sessile, pentamerous, the calyx vestigial, eventually forming a hardly visible wavy ring, ca. 0.05 × 0.3 mm. Staminate flowers white to cream, campanulate, 3.3 × 2.7 mm, corolla tube 1.2–1.4 mm long, glabrous, gibbous 0.5–0.7 mm length; lobes 5(6), elliptical, 1.2 × 1 mm, acute apex; stamens 3(4), 2.8–3 mm long, exserted, epipetalous, adnate to the corolla in the basal third, filament 2.1–2.3 mm long, the anthers globose to oblong, 0.7–0.8 × 0.6 mm, dorsifixed; pistilodium 1.6–1.8 mm long, slightly exserted, stigma 3 with linear lobes, up to 0.3 mm long; abortive ovary (partially developed) 1–1.6 mm long. Pistillate flowers white to cream, campanulate, 1.3–1.6 × 1.3–1.4 mm, corolla tube 0.7–0.8 mm long, glabrous, with a small gibbosity of 0.3–0.5 × 0.1–0.2 mm; lobes 5 (6), acute to rounded, 0.6–0.8 × 0.4–0.5 mm, slightly unequal; vestigial staminodes 3, 0.3–0.5 mm long, epipetalous, included and inconspicuous, laminar and apically spatulate to clavate; style 1.6 mm long, exserted; stigma 3, papillose, rounded lobes, 0.1 mm long; ovary 0.7 × 0.5 mm. Fruits elliptic, 2.4–3.4 × 1.5–2.5 mm, apex emarginate to valeculate, base rounded to truncate, eventually cordate, transversely triquetrous, 3–5-costate (with two lateral and one or three adaxial vascular bundle, 0.2–0.3 mm wide), two prominent lateral wings, 0.3–0.8 mm wide, dorsal costa 0.2–0.3 mm wide, slightly oblique, stramineous, glabrous, finely papillose, uniseminate. Seeds elliptical, rounded apex and base, 1.5 × 0.8–1 mm, stramineous, smooth indument.
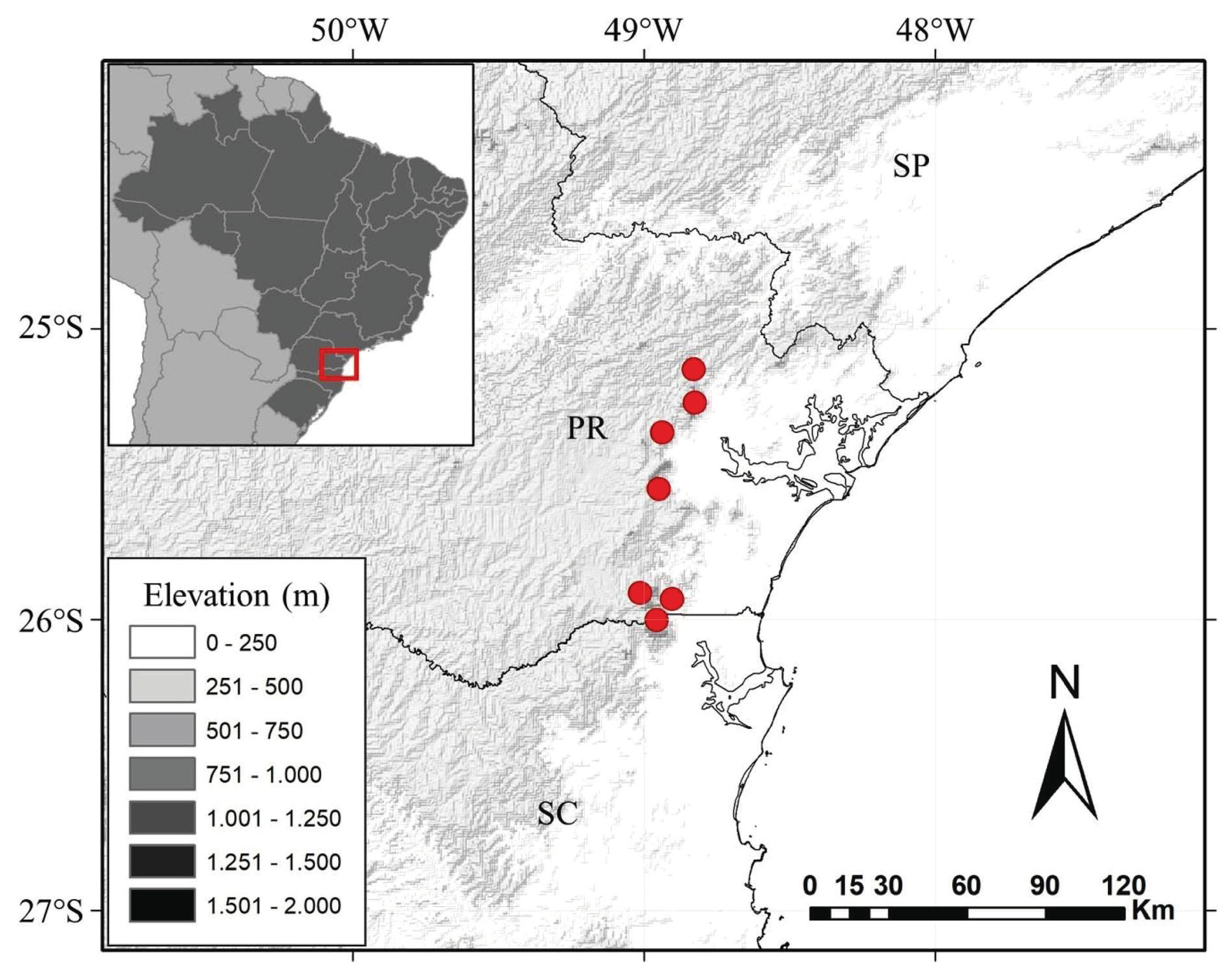
Distribution and Habitat: — Valeriana sobraliana is endemic from the High Altitude Tropical Grasslands of Paraná state and border region with Santa Catarina state, Southern Brazil. Sparse populations were sampled in highland grasslands and montane cloud forest edges, on humid slopes, rocky outcrops and mountain peaks, environments often covered by fog, from 1100 to 1950 m elevation ( Fig. 3 View FIGURE 3 ).
Conservation status:— According to the IUCN (2017) criteria B2ab (iii, iv), the species is classified as Vulnerable ( VU). Despite the great number of collections deposited in regional herbaria and the wider extent of occurrence ( EOO = 1,120.000 km²), V. sobraliana has less than ten subpopulations sampled and reduced area of occupation ( AOO = 2000 km ²). It occurs at least in four Conservation Units: Guaricana National Park, Pico do Paraná State Park, Pico do Marumbi State Park and Guaratuba Environmental Protection Area. The main anthropic disturbances in the habitats where the new species occurs are the coverage by Pinus spp. , plantations and exotic grasses, which often invade natural areas (including Conservation Units), as well as the high incidence of uncontrolled fire, leading to less than ten locations.
Etymology: —The new species is named in honor of the Brazilian botanist Marcos Sobral, great researcher and collector of Valerianaceae in Southern Brazil, the first to mention the morphological specificities of the material here analyzed ( Sobral 1999a: 25).
Phenology: —Flowers and fruits from February to December.
Paratypes: — BRAZIL. Santa Catarina: Campo Alegre, Serra do Quiriri , 29 December 1998 (pist. fl. and fr.) J.M.Silva, O.S.Ribas, J.Cordeiro & E.Barbosa 2782 ( MBM!) ; Campo Alegre, Serra Quiriri , 1400 m elev., 29 September 2001 (stam. fl.) O.S.Ribas, J.M.Silva, E.Barbosa & E.F.Costa 3642 ( MBM!) . Garuva, Serra do Quiriri , 1300 m elev., 16 December 2004 (stam. fl.) J.M.Silva, O.S.Ribas & M.R.Bornschein 4149 ( MBM!) ; Garuva, Serra Quiriri , 1400 m elev., 26 August 2010 (stam. fl.) E.F.Costa, E.Barbosa & J.Cordeiro 104 ( MBM!) . Paraná: Campina Grande do Sul, Serra do Capivary Grande , 1700 m elev., 6 August 1961 (pist. fl.) G.Hatschbach 8175 ( MBM!) ; Campina Grande do Sul, Pico Caratuva , 1950 m elev., 20 May 1967 (pist. fl.) G.Hatschbach 16460 ( MBM!, LIL) ; Campina Grande do Sul, Pico Caratuva , 2 August 1967 (stam. fl.) G.Hatschbach 16842 ( MBM!) ; Campina Grande do Sul, Serra Ibitiraquire , Abrigo 1, 1600 m elev., 25 September 1969, G.Hatschbach 22222 ( MBM!, HBR!) ; Campina Grande do Sul, Pico Paraná , 1600 m elev., 2 April 1988, M.R.Bornschein 16 ( MBM!) ; Campina Grande do Sul, Serra do Capivari , 24 August 1989 (stam. fl.) V.Nikolack & O.S.Ribas 10 ( FLOR!, MBM!) ; Campina Grande do Sul, Serra dos Órgãos , Pico Caratuva , 1550 m elev., 4 July 1991 (stam. fl.) Y.S.Kuniyoshi & A.Vicentini 5424-b ( MBM!) ; Campina Grande do Sul, Serra do Ibitiraquire , subida para o Pico Paraná , 1500–1700 m elev., 14 July 1996 (stam. fl.) O.S.Ribas & F.Schwerdt 1456 ( MBM!) ; Campina Grande do Sul, Morro Capivari Grande , 1660 m elev., 1 July 1998 (stam. fl.) V.A.O.Dittrich 393 ( MBM!) ; Campina Grande do Sul, Serra do Ibitiraquire , Pico Itapiroca , 1800 m elev., 9 August 2000 (stam. fl.) M.Scheer & A.Y.Mocochinski 454 ( MBM!) ; Campina Grande do Sul, Serra do Ibitiraquire , Pico Paraná , Abrigo 2, 1700 m elev., 18 August 2001 (stam. fl.) J.M.Silva, E.Barbosa, O.S.Ribas & E.F.Costa 3386 ( MBM!) ; Campina Grande do Sul, Serra Capivari Grande , 23 October 2001 (pist. fl. and im. fr.) E.Barboza, O.S.Ribas & E.F.Costa 681 ( MBM! FURB!) ; Campina Grande do Sul, Morro Capivari , 21 October 2006 (pist. fl. and fr.) A.L.Gasper 249 ( FURB!) ; Campina Grande do Sul, Pico Caratuva , 5 August 2009 (pist. fl.) E.D.Lozano 119 ( MBM!) ; Campina Grande do Sul, Morro Cerro Verde , 5 June 2010 (pist. fl. and fr.) E.D.Lozano & V.Ariati 249 ( MBM!) ; Campina Grande do Sul, lateral do Caratuva , trilha para o Pico Paraná , 4 July 2010 (stam. fl.) R.Ristow 713 ( MBM!) ; Campina Grande do Sul, Abrigo II , Pico do Paraná , 10 July 2011, (pist. fl. and fr.) E.D.Lozano, T.Bochorny, G.Felitto & D.N.Takigawa 617 ( MBM!) ; Campina Grande do Sul, Pico Caratuva , 25°14’26.6”S 48°49’50.8”W, 1767 m elev., 11 September 2018 (stam. fl.) C.Rabuske-Silva, G.P.Coelho & C.L.Ribeiro 349 ( ICN!) GoogleMaps . Guaratuba, Serra de Araçatuba , Morro dos Perdidos , 1100 m elev., 18 September 1997 (stam. fl.) E.P.Santos & H.M.Fernandes 346 ( MBM!) ; Guaratuba, Serra do Araçatuba , 1300 m elev., 25 February 2000 (pist. fl. and fr.) J.M.Silva, E.Barbosa & J.Cordeiro 3256 ( ICN!, MBM) ; Guaratuba, Pico Pedra Branca de Araraquara , 25°56’23”S 48°52’22”W, 1160 m elev., 12 August 2014 (pist. fl. and fr.) J.Cordeiro, E.Barbosa & M.L.Brotto 5229 ( MBM!) GoogleMaps . Morretes, Pico Olimpo , Serra Marumbi , 25 August 1946 (stam. fl.) O.Curial 29 ( MBM!) ; Morretes, Morro Mãe Catira , Serra Graciosa , 30 March 1947 (stam. fl.) O.Curial 152 ( MBM!) ; Morretes, Morro Mãe Catira , Serra Graciosa , 30 March 1947 (pist. fl. and fr.) O.Curial 153 ( MBM!) . Piraquara , 23 September 1970 (pist. fl.) H.Imaguire 2579 ( ICN!) . Quatro Barras, Morro Mãe Catira , 1200 m elev., 3 August 1988 (stam. fl.) R.Kummrov & C.Budziak 3039 ( MBM!) ; Quatro Barras, Morro Mãe Catira , 17 August 1989 (stam. fl.) R.Kummrow, V.Nicolack et al. 3168 ( MBM!, HUCS!) ; Quatro Barras, Morro Mãe Catira , 900 m elev., 10 July 1991 (stam. fl.) O.S.Ribas & C.B.Poliquesi 335 ( MBM!) ; Quatro Barras, Serra do Capivari Grande , 1600 m elev., 24 October 1997 (stam. fl. and fr.) E.P.Santos, A.C.Cervi, C.M.S.Coimbra & E.F.Armando 405 ( MBM!, UPCB) ; Quatro Barras, trilhas morros Mãe Catira e Sete , 3 August 2013 (stam. fl.) M.E.Engels, E.D.Lozano, M.Bolson, T.Bochorny & L.Bacci 1410 ( MBM!) . São José dos Pinhais, Serra da Igreja , Morro dos Padres , 1320 m elev., 6 August 2001, M.Scheer, A.Cavassani & G. Gatti 362 ( MBM!) . Tijucas do Sul, Morro dos Perdidos , 25°53’27.3”S 48°57’23.9”W, 1420 m elev., 13 September 2018 (stam. and pist. fl.) C.Rabuske-Silva, G.P.Coelho & C.L.Ribeiro 352 ( ICN!) GoogleMaps .
Comments: — Borsini (1962: 159; 1963: 128) based the analysis, description and illustration of Valeriana ulei on material coming exclusively from Paraná state, Southern Brazil. Sobral (1999a: 25) observed that the specimens examined by Borsini had quite different features from those mentioned in the protologue of V. ulei , being possibly a new species for Science. The analysis of both protologue and type specimen ( HBG barcode HBG-513375, image!) confirmed Sobral’s observation. The new species is here described based on its distinct morphology and geographical concision.
Regarding vegetative traits, all four morphologically related species are subshrubs and the most basal caulinar leaves may be grouped into subrosettes ( Figure 3 View FIGURE 3 , B and 4 View FIGURE 4 , A, C and E). Despite these similarities, the leaves have some useful characters for species differentiation. Valeriana sobraliana has leaf blades strongly discolorous, with a dark green and waxy adaxial face, and markedly light-green abaxial face. Plus, it is pinnately veined, the veins anastomosing and not reticulate, with a chartaceous or at most slightly coriaceous consistency and glabrous indument ( Figure 3 View FIGURE 3 , C and D). On the other hand, V. catharinensis , although sharing the same type of venation and margins, has slightly discolorous, chartaceous and pubescent leaves ( Figure 4 View FIGURE 4 , A and B). Valeriana organensis shares the same leaves consistency and indument with V. sobraliana , but its leaves are slightly discolorous, the venation is anastomosing and reticulate and the margins is serrulate to serrate ( Figure 4 View FIGURE 4 , C and D). Valeriana ulei shares the glabrous indument, although the consistency is strongly coriaceous, and only the primary vein and the first or second pair of secondary veins are visible, the first pair forming a more narrower angle (about 30°) than the others and reaching the apical third of the leaf blade (3-veined-like), slightly anastomosing ( Figure 4 View FIGURE 4 , E and F).
The examined specimens indicate that V. sobraliana is a dioecious species, despite the well developed pistilodium observed in staminate flowers. The occurrence of partially developed ovaries, aborted during the anthesis and similar to thickened floral peduncles, is common in staminate flowers of the genus ( Borsini 1963). No seeds were observed in these structures, which corroborates this classification. The achenes exhibit considerable variation in shape according to the stage of development. Usually, in the final stage, they acquire the elliptic shape. The lateral wings protrude apically, resulting in an emarginate to valeculate apex ( Figure 1 View FIGURE 1 , G and H).
Valeriana sobraliana does not occur in sympatry with other species of the genus and presents a geographic disjunction with the morphologically related species. Valeriana catharinensis and V.ulei are restricted to the Subtropical Highland Grasslands ( SHG) and Subtropical Mixed Forest ( SMF) of the Serra Geral plateau (sensu Iganci et al. 2011), which has basaltic rocks. The distribution of V. sobraliana corresponds to the exact distribution of the High Altitude Tropical Grasslands ( HATG) in the Serra do Mar mountain range of Paraná and border region with Santa Catarina state, between the massifs of the Serra do Quiriri (or Serra do Iqueririm) and the Serra Capivari Grande (south and north respectively) ( Maack 1968). Besides, the HATG covers basically granitic rocks. Valeriana organensis , despite also occurring in the HATG, is restricted to the states of São Paulo, Rio de Janeiro and Minas Gerais, Southeastern Brazil, occurring in both the Serra do Mar and the Serra da Mantiqueira mountain ranges.
Checklist of Valeriana in the state of Paraná: — Valeriana shows a decrease in the number of species from Southern to Southeastern Brazil ( Rabuske-Silva 2018). Rio Grande do Sul is the state with the highest number of species (12 spp.), followed by Santa Catarina (10 spp.) and Paraná (5 spp., Table 2 View TABLE 2 ). In Paraná, three species are threatened according to the IUCN categories and criteria, all of them Vulnerable ( VU): V. eichleriana (C.A.Müll.) Graebn. , V. reitziana Borsini and V. sobraliana ( Rabuske-Silva 2018) . This is basically due to the populations of these species being naturally sparse and the conversion of the remaining grasslands for agriculture, livestock and urban occupation. Valeriana eichleriana , V. reitziana and V. salicariifolia Vahl are restricted to the SHG, and V. sobraliana is the only species restricted to the HATG. Valeriana scandens L., a voluble or clambering species, has the widest distribution, with scattered records in all the state. Valeriana eichleriana and V. reitziana , two morphologically closely related subshrubs, reach their northern limit of distribution in the SHG of Paraná. Valeriana eichleriana is the rarest species in the study area, with only one record from the municipality of Palmas (S.Campestrini, R.Trevisan, J.P.Ferreira & S.Venturi 618, FLOR!), on the border with Santa Catarina, parallel 26°S. The populations of this species are concentrated in the Aparados da Serra region, in Rio Grande do Sul, being also one of the rarest species in Santa Catarina, with few historical records in the municipality of Lages. Although the type material of Valeriana reitziana was collected in the Serra do Caracol, Minas Gerais state, Southeastern Brazil, this species is considered extinct in this state and is currently restricted to Southern Brazil ( Rabuske-Silva 2018). Valeriana reitziana reaches its northern distribution limit in the center of the Paraná state, also occurring in the municipality of Palmas, plus in the municipalities of Guarapuava, Laranjeiras do Sul and Palmeira, until the parallel 25°S. Valeriana salicariifolia , a species that occurs from northeastern Argentina, restricted to swamps and streams, reaches its northern distribution limit in the state of São Paulo, in the municipality of Itapeva, near the border with Paraná, parallel 23°S ( Kutschker 2008, Scalon et al. 2002, Rabuske-Silva 2018).


| MBM |
Myanmar, Yangon, Hlawga Park, Forest Department, Biodiversity Museum |
| FLOR |
FLOR |
| FURB |
FURB |
| ICN |
Colombia, Bogota, Universidad Nacional de Colombia, Insituto de Ciencias Naturales de la Universidad Nacional |
| LIL |
LIL |
| HBR |
HBR |
| HUCS |
HUCS |
| UPCB |
UPCB |
| HBG |
HBG |
| SHG |
SHG |
| SMF |
Germany, Frankfurt-am-Main, Forschungsinstitut und Naturmuseum Senckenberg |
| HATG |
HATG |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |