
Tropidurus azurduyae, Carvalho & Rivas & Céspedes & Rodrigues, 2018
|
publication ID |
https://doi.org/ 10.1206/3896.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03C087EC-FF87-FFFC-FE77-1B90B4CCF93D |
|
treatment provided by |
Carolina |
|
scientific name |
Tropidurus azurduyae |
| status |
sp. nov. |
Tropidurus azurduyae , n. sp.
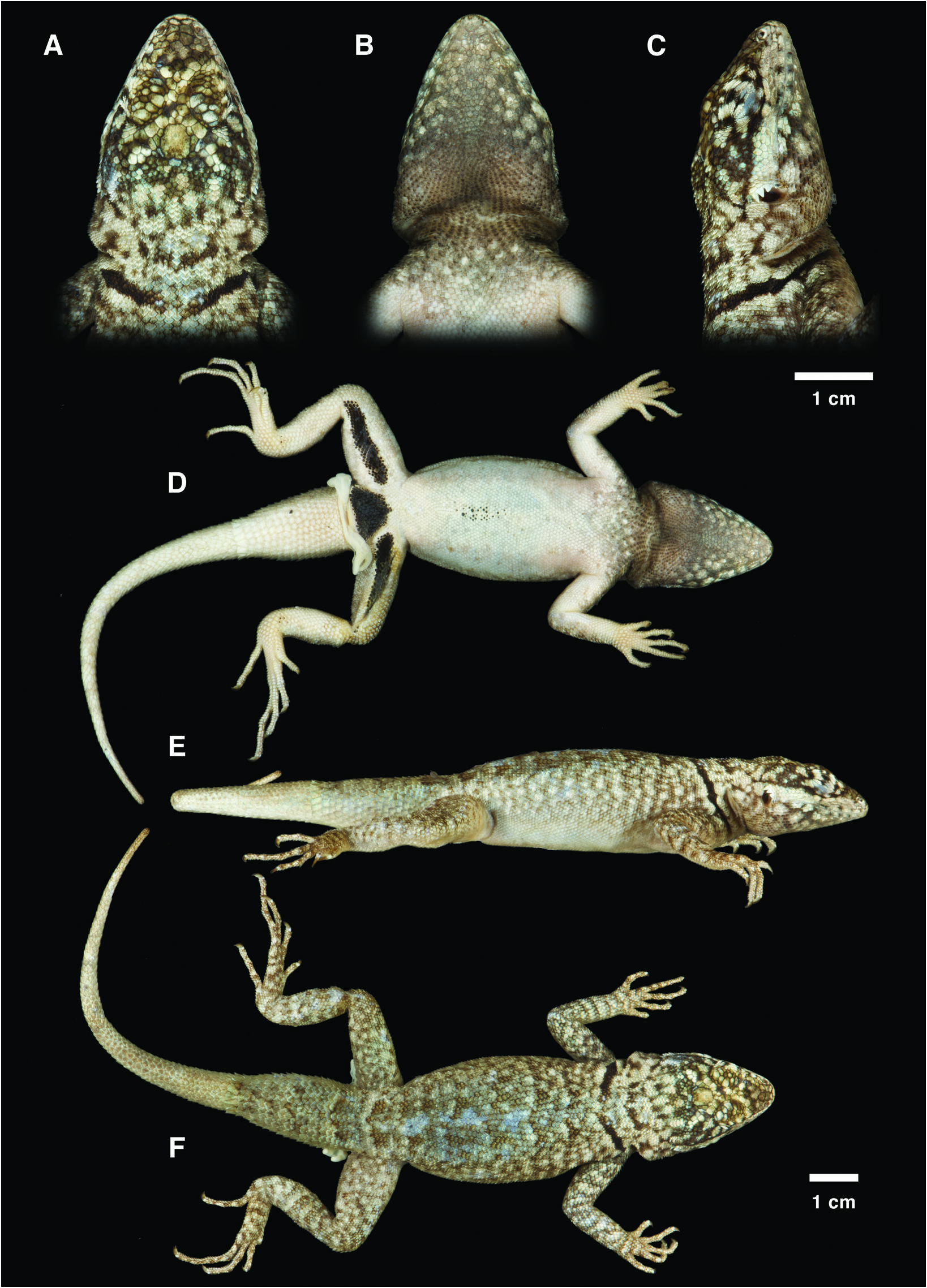
Figures 1H View FIGURE 1 , 3E–H View FIGURE 3 , 4A–F View FIGURE 4
HOLOTYPE: MHNC-R 3011 , adult male from Parque Nacional Torotoro , Potosí, Bolivia (18° 5′ 54.24″ S, 65° 44′ 57.48″ W — WGS84 system; ~ 2264 m), collected by A.L.G. Carvalho, M.A. Sena, L. R. Rivas, G. Juchazara, E. Lujo, and F. Mamani in 13 November 2013. GoogleMaps
ALLOTYPE: MHNC-R 3009 , adult female, same locality as holotype (18° 5′ 53.88″ S, 65° 44′ 57.12″ W — WGS84 system; ~ 2262 m), collected by A.L.G. Carvalho, M.A. Sena, L. R. Rivas, G. Juchazara, E. Lujo, and F. Mamani in 13 November 2013. GoogleMaps
PARATYPES: MHNC-R 3007 , adult female, same locality as holotype (18° 6′ 15.48″ S, 65° 45′ 36.00″ W — WGS84 system; ~2569), collected by A.L.G. Carvalho, M.A. Sena, L GoogleMaps . R. Rivas, J. Choque, J. Kamaqui, and E. Lujo in 14 November 2013 . MHNC-R 3008 , adult female, same locality (18° 6′ 25.56″ S, 65° 45′ 27.36″ W — WGS84 system; ~ 2579 m), collected by A.L.G. Carvalho, M.A. Sena, L GoogleMaps . R. Rivas, J. Choque, J. Kamaqui, and E. Lujo in 14 November 2013 . MHNC-R 3010 , adult male, same locality (18° 5′ 52.32″ S, 65° 44′ 57.48″ W — WGS84 system; ~ 2269 m), collected by A.L.G. Carvalho, M.A. Sena, L GoogleMaps . R. Rivas, G. Juchazara, E. Lujo, and F. Mamani in 13 November 2013 . MHNC-R 3012 , adult male, same locality (18° 6′ 25.56″ S, 65° 45′ 25.92″ W — WGS84 system; ~ 2566 m), collected by A.L.G. Carvalho, M.A. Sena, L GoogleMaps . R. Rivas, J. Choque, J. Kamaqui, and E. Lujo in 14 November 2013 . MHNC-R 3015 , adult male, same locality (18° 6′ 25.92″ S, 65° 45′ 25.92″ W — WGS84 system; ~ 2562 m), collected by A.L.G. Carvalho, M.A. Sena, L GoogleMaps . R. Rivas, J. Choque, J. Kamaqui, and E. Lujo in 14 November 2013 . MHNC-R 3016 , adult male, same locality (18° 5′ 54.24″ S, 65° 44′ 55.32″ W — WGS84 GoogleMaps
70° 68° 66° 64° 62° 60° 58°
system; ~ 2556 m), collected by A.L.G. Carvalho, M.A. Sena, L. R. Rivas, G. Juchazara, E. Lujo, and F. Mamani in 13 November 2013 . MHNC-R 3017 , adult male, same locality (18° 5′ 55.32″ S, 65° 44′ 58.56″ W — WGS84 system; ~ 2274 m), collected by A.L.G. Carvalho, M.A. Sena, L GoogleMaps . R. Rivas, G. Juchazara, E. Lujo, and F. Mamani in 13 November 2013 . MHNC-R 3020 , adult male, same locality (18° 5′ 11.88″ S, 65° 45′ 44.28″ W — WGS84 system; ~ 2596 m), collected by A.L.G. Carvalho, M.A. Sena, L GoogleMaps . R. Rivas, J. Choque, J. Kamaqui, and E. Lujo in 14 November 2013 . MHNC-R 3024 , juvenile female, same locality (18° 5′ 55.32″ S, 65° 44′ 57.48″ W — WGS84 system; ~ 2269 m), collected by A.L.G. Carvalho, M.A. Sena, L GoogleMaps . R. Rivas, G. Juchazara, E. Lujo, and F. Mamani in 13 November 2013 . MHNC-R 3026 , juvenile female, same locality (18° 5′ 54.24″ S, 65° 44′ 55.32″ W — WGS84 system; ~ 2256 m), collected by A.L.G. Carvalho, M.A. Sena, L GoogleMaps . R. Rivas, G. Juchazara, E. Lujo, and F. Mamani in 13 November 2013 .
MORPHOLOGICAL DIAGNOSIS: Tropidurus azurduyae is here morphologically diagnosed as a Tropidurus based on the observation of a set of characters suggested by Frost et al. (2001) as exclusive to the genus: skull not highly elevated at the level of the orbits; “flash” marks on underside of thighs present; circumorbitals distinct from other small supraorbital scales; lateral fringe not developed on both sides of fourth toes; enlarged middorsal scale row absent; tail terete; and hemipenis attenuate without apical disks. The presence of a maxilla not broad, nutritive foramina of maxilla strikingly enlarged, lingual process of dentary extending over lingual dentary process of coronoid, angular strongly reduced, and absence of medial centrale could not be examined without dissecting or clearing and staining specimens. These characters should be revised whenever larger series of individuals become available.
Tropidurus azurduyae is a member of the T. torquatus group per Frost et al. (2001). It differs from other species groups by lacking an enlarged middorsal scale row (well marked in species of the T. spinulosus group, especially in males), by exhibiting black “flash” marks on the underside of thighs and cloacal flap of adult males (yellow, cream, or orangey “flash” marks are present in males of the T. spinulosus group), and also by lacking a dorsoventrally flattened body (as observed in species of the T. semitaeniatus group and, more moderately, in T. bogerti ).
Tropidurus azurduyae is the only species in the genus with lower flanks pigmented orange, a condition consistently observed in both sexes (fig. 3E–H). Its ventral head is darkly pigmented and offers contrast to the light circular blotches present on chin and also laterally (fig. 3G, H). The ground color of its throat is charcoal gray impregnated with strong orange coloration (fig. 3G, H). A pair of mite pockets is present on the lateral neck, with the posterior one larger; the anterior pocket originates lower than the posterior, but both usually end ventrally at the same level (fig. 3F). No pockets are found in the armpit and inguinal region of the new species. An elliptical or subrhomboidal black mark is present on the mid venter of adult males of T. azurduyae in addition to black “flash” marks on the underside of thighs and precloacal flap (fig. 3H). Tropidurus azurduyae is saxicolous, but may climb tree trunks and fallen logs occasionally (fig. 1H). In combination, this set of characters provides a safe diagnosis, distinguishing T. azurduyae from all other congeners.
COMPARISON WITH OTHER SPECIES: Tropidurus azurduyae , T. cocorobensis , T. chromatops , T. etheridgei , T. hygomi , and T. psammonastes are the only species of the T. torquatus group that have two mite pockets on the lateral neck and lack differentiated skin folds or pockets in the axillary and inguinal regions. Although T. catalanensis , T. imbituba , and T. torquatus also have two mite pockets on the lateral neck, all three species exhibit 2–3 shallow granular areas in the axillary region and a fully developed granular inguinal pocket. The two pockets on the lateral neck of T. azurduyae are not exceptionally broadened nor deep, and they differ from the extremely enlarged lateral neck pockets of T. chromatops (fig. 5). They are also slightly distinct from the pockets of T. hygomi , which are oblique and deep, with the anterior one positioned more ventrally than the posterior one in both species. Tropidurus hygomi and T. azurduyae can be further distinguished by the presence of expanded scales covering the supraocular area of the former species. The anterior lateral neck pocket of T. azurduyae is coated with granular scales, while this same structure in T. psammonastes is coated with regular scales (only the posterior one is granular). For a more comprehensive summary of mite-pocket morphologies and their taxonomic distribution in the T. torquatus group, refer to Rodrigues (1987: figs. 1–13) and Carvalho et al. (2016: table 2, fig. 8).
Tropidurus azurduyae is saxicolous, and its ecology contrasts markedly with the psammophilous habit of three other species of the T. torquatus group with two lateral neck mite pockets, T. cocorobensis , T. hygomi , and T. psammonastes . It can also be distinguished from T. cocorobensis and T. hygomi based on its larger body size (SVL: 66.06–104.85 mm in males and 62.83–89.58 mm in females of T. azurduyae , 61.39–74.59 mm in males and 60.27–65.37 mm in females of T. cocorobensis , and 54.15–67.30 mm in males and 48.08–60.38 mm in females of T. hygomi ). The new species also lacks the 2–4 well-marked black ocellar spots that decorate the upper flanks of T. cocorobensis anteriorly, from nuchal collar, just above the humerus, reaching to the middle of the body. Tropidurus azurduyae differs from T. chromatops in terms of coloration by lacking an intense burnt-red dorsal head and a facial mask with touches of blue and cream (figs. 5, 6A–D). The new species exhibits a champagne background, mottled with dark grayish-brown and lead pigmentation, and dark ventral head. This coloration is fairly distinct from the dirty-yellow dorsal background decorated with a brown reticulated pattern, and light ventral head ornate with a loose reticulum or semireticulum, found in T. etheridgei (pattern better marked in males than females; fig. 3A–D). Moreover, with regard to coloration, the lower flanks and gular region pigmented in orange in both sexes is, to our knowledge, exclusive to T. azurduyae .
DESCRIPTION OF HOLOTYPE (figs. 3F, H, 4A–F): Medium-sized specimen of Tropidurus, SVL 87.31 mm; head triangular, length 30% of SVL and width 71% of head length; skull not compressed, not strongly elevated at level of orbits; rostrum not noticeably shortened relative to most other species in the genus; scales of frontonasal region not imbricating posteriorly, lenticulate scale organs distributed on the head, more abundant on the frontonasal and supraocular areas; rostral tall, about 3× (in lateral view) as high as first supralabial, contacting first supralabials, first lorilabials, nasals, and two postrostrals; 1/1 postrostrals; nasal single, slightly protruding, pentagonal, elongated anteroposteriorly with the tip of the pentagon directed anteriorly, in contact with rostral; 6/7 enlarged supralabials followed by 3/6 smaller scales reaching the rictus oris, never contacting subocular; nostril elliptical, occupying about 1/3 of nasal, positioned posteriorly, directed posterolaterally; 3/3 canthals; anteriormost canthal separated from supralabials by 1/1 rows of lorilabials; 8/8 laminate superciliary scales weakly produced vertically; 1/1 dorsally keeled preoculars contacting third canthal and 3/3 loreals; 2/1 suboculars dorsally keeled, elongate, separated from supralabials by one row of lorilabials posteriorly; palpebrals granular; second row of palpebrals larger, with scale organ on tip, central palpebrals unpigmented, nearly translucent; pupil circular; 3/3 main rows of supraoculars, oblique internal row with 8/8, medial row with 8/8, external row with 7/7 scales, the enlarged internal ones occupying up to half the width of the supraocular area; 1/1 rows of small, angulate circumorbitals; 1/1 rows of short semilaminate scales separating circumorbitals from superciliaries; interparietal enlarged, about 1.2× longer than wide; parietal eye visible, positioned medially on the posterior limit of the first third of the interparietal scale; temporals slightly imbricate, keeled, at least 3× larger than lateral neck scales and smaller than dorsals and parietals; ear shaped like inverted keyhole, canal deep, largest diameter (~ 5.5 mm) of ear opening 25% of ear opening to snout distance; tympanum translucent; preauricular fringe consisting of row of 6/6 smooth, lanceolate scales; width of mental 60% of the width of rostral; mental extending posteriorly to the level of half of the first adjacent infralabials; 7/7 enlarged infralabials followed by 3/3 smaller scales reaching the rictus oris; 4/4 angulate, enlarged postmentals; 1/1 postmentals in contact with first infralabial; first postmentals not in contact; 11/11 sublabials; 46 gulars, imbricating posteriorly.
Vertebral crest absent; 85 dorsals; 82 scale rows around midbody; 79 ventrals; dorsals large, strongly keeled and mucronate, particularly on the dorsal neck; keels on dorsal and caudal scales align forming continuous, longitudinal, slightly oblique lines observable macroscopically; postumeral region with small, nearly granular, smooth, nonmucronate scales, increasing in size, intensity of keels and mucronation toward the flanks; ventrals smooth, nonmucronate, imbricate, about half the size of dorsals; midventral, dark-pigmented “flash” mark present, not intensely marked, subrhomboidal; “flash” marks on underside of thighs formed by 6/6 rows of dark glandular scales; 13 cloacal scales, cloacal flap with 12 rows of dark precloacal glandular scales; supracarpal scales smooth near finger I and slightly keeled toward finger V, rhomboidal or subrhomboidal; supratarsal scales smooth toward finger I and keeled and mucronate toward finger V, rhomboidal; both supracarpals and supratarsals with very rare scale organ positioned on the distal end of the scale, when present; infracarpal and infratarsal scales carinate, tricarinate toward fingers and toes; fingers and toes thin, cylindrical, slightly compressed laterally; supradigital lamellae keeled, rhomboidal, scale organ positioned on the distal end of the scales, when present; infradigital lamellae tricarinate and mucronate, 17/17 under fourth finger + ungual, 26/24 under fourth toe + ungual, medial careen larger and more projected than laterals; claws long, curved; preaxial scales of forearm strongly keeled and mucronate grading to smooth scales with no or short mucrons and smaller size on ventral and postaxial surfaces; 28/28 tibial scales, keeled and mucronate; dorsal body scales large, keeled, mucronate; lateral neck scales several times smaller than dorsals; rictal, nuchal, postauricular, supraauricular, dorsolateral, and antegular fold absent; shallow postauricular depression present; oblique neck fold well marked defining two lateral neck mite pockets on each side of the neck; anteriormost mite pocket half the size and originating lower than the posteriormost, both ending ventrally at the same level; antehumeral fold present and well marked, coated with imbricate scales similar to those on lateral neck; gular fold incomplete medially; axillary and inguinal mite pockets absent; tail slightly compressed laterally, regrown, tapering from the end of the first third to a point; caudal verticils absent; scales of tail imbricate, keeled, mucronate, up to 3× larger than dorsals.
COLORATION IN LIFE (fig. 3F, H): Dorsal head with champagne background, mottled with brownish and lead pigmentation. A champagne facial stripe decorates the lateral head, covering labials, lorilabials, loreals, inferior portion of preocular and suboculars, lower temporals, and preauricular fringe. Keeled portion of preocular and suboculars, palpebrals, superciliaries, and upper temporals dark pigmented, similar in coloration to dorsal head. Iris golden brown. Mental region champagne grading into a lead ventral head with touches of champagne. Sublabial and posterolateral region of the head decorated with champagne blotches, 1–7 scales in size, that reach the area behind ear opening. Throat charcoal gray with intense orange pigmentation permeating its dark background until the antegular fold. Area between antegular fold and beginning of chest mottled with lead pigmentation and touches of yellow. Neck and dorsal body champagne, mottled with brownish and lead pigmentation; dark coloration more concentrated along the vertebral area. Nuchal collar black, well marked, nearly complete dorsally, formed by 4–5 rows of dark scales extending from humeral attachment to the vertebral area, outlined by 2–4 rows of champagne scales. Uppermost limits of flanks similar to dorsum, lower flanks pigmented in orange from axillary to inguinal region. Chest cream, anteriorly sprinkled with lead pigmentation. Ventral ground coloration cream; black spotted subrhomboidal mark oriented anteroposteriorly present on the mid venter. Limbs with champagne background and lead pigmentation forming a pattern similar to dorsum anteriorly (arms and thighs), and a slightly diffuse, stripelike ornamentation perpendicular to limb axis posteriorly, including forearms and legs and supracarpal and supratarsal regions, and digits. Femoral and precloacal “flash” marks well marked, black in color, 31/33 and 12 scales long, respectively. Tail greenish champagne, mottled with lead pigmentation anteriorly; regenerated section greenish champagne; ventral side of the whole tail pale cream.
COLORATION IN PRESERVATIVE (fig. 4A–F): Overall coloration pattern of head and body preserved. Champagne background partially faded into pale cream and lead pigmentation became lighter, gaining a brownish tone. Facial stripe decorating the lateral head preserved. Same is true for the light blotches distributed over sublabial and posterolateral area of the head, and area behind ear opening. Originally lead-pigmented area of ventral head became brownish. Intense orange pigmentation of gular region faded almost completely, remaining a merely elusive orangey tone over brownish scales. Nuchal collar remained well marked. Orange coloration on the lower flanks was completely washed out, increasing the contrast between the now paler background and brownish mottled pigmentation that decorates the lateral body. Loss of orange pigmentation revealed irregular light blotches composing the mottled pattern on the flanks. Lead pigmentation sprinkled anteriorly on the chest became brownish. Venter preserved its cream aspect. Dark “flash” marks fully preserved underneath the thighs, cloacal flap, and mid venter. Limbs gained a slightly lighter background and brownish pigmentation in the place of original lead tones. Tail coloration nearly unaltered.
MEASUREMENTS OF HOLOTYPE (in mm): SVL 87.31 , TL 96.08 (regrown), HH 13.02, EOS 22.11 , HL 25.99, HW 18.58 , AL 15.80, FAL 13.36 , HDL 15.94, THL 21.16 , SL 17.72, FOL 29.72 , AGD 36.78.
MORPHOMETRICS: Tropidurus azurduyae is a middle-sized species of the T. torquatus group, with adult males ranging from 66.06 to 104.85 mm SVL and females from 62.83 to 89.58 mm SVL. This species is statistically indistinct in body size from its closest, formally described relatives, T. chromatops and T. etheridgei , but males of T. chromatops were found to be (marginally) larger than those of T. etheridgei (ANOVA SVL: males: df (degrees of freedom) = 2, sum of squares = 0.017, mean square = 0.008, F value = 3.563, p = 0.037; females: df = 2, sum of squares = 0.006, mean square = 0.003, F value = 1.668, p = 0.200; tables 2–3). In terms of shape, PCA captured extensive overlap among species and showed relatively similar contribution of most variables for morphometric groupings, while LDA showed better success discriminating groups (fig. 7; table 4). For males, LDA indicated FAL and EOS as important variables separating species in LD1, and EOS, FAL, and THL in LD2. For females, FAL and AL contributed more for species discrimination in LD1, and FOL, HW, and HL in LD2. LDA functions reached>85% correct reclassifications, distinguishing male individuals of all three species and female individuals of T. etheridgei and T. azurduyae (table 5). However, correct reclassifications dropped considerably with the implementation of the leave-one-out cross-validation procedure, indicating that morphometric parameters alone may not safely distinguish all species analyzed (table 5).
MERISTICS: Tropidurus chromatops , T. etheridgei , and T. azurduyae overlap at least partially in most scale counts (figs. 8–10; table 6). Tropidurus etheridgei has, in general, lower scale counts in comparison to the other species analyzed; exceptions were observed only in the number of subdigital lamellae (fig. 9). Number of gulars and scales around midbody differ between T. chromatops and T. etheridgei (males only), but cannot be used to fully distinguish any of these species from T. azurduyae (fig. 8; table 6). Although a few individuals overlap (fig. 9), T. azurduyae has in average a higher number of tibials than T. chromatops and T. etheridgei (table 6). PCA based on meristic variables showed complete separation between T. azurduyae and T. etheridgei , but not in relation to T. chromatops (fig. 10; table 7). Number of tibials provided the strongest contribution for species groupings in both PCA and LDA, followed by the number of gulars, ventrals, and subdigital lamellae (table 7). LDA effectively discriminated species, showing correct reclassification>90% in all cases but one (female T. chromatops ). Even with the implementation of the leave-one-out cross-validation procedure, correct reclassifications remained high; T. chromatops was the only poorly discriminated species (table 7). In general, scale counts can be used to distinguish T. azurduyae from T. etheridgei , and this latter species from T. chromatops , but they may be insufficient to separate T. azurduyae from T. chromatops (though the number of tibials is informative in most cases). For specimens with overlapping scale counts, additional diagnostic characters treated in Comparisons with Other Species should be considered.
ETYMOLOGY: The species name azurduyae is a noun in the feminine genitive case honoring Juana Azurduy de Padilla (Chuquisaca, Bolivia: July 12, 1780 – May 25, 1862), one of the most distinguished Latin American leaders who bravely fought for the independence of the Spanish territory of Upper Peru, which comprised part of today’s Bolivia and Peru, and formed along with Argentina, Uruguay, and Paraguay the Viceroyalty of the Río de La Plata during colonial times. Her memory remained nearly forgotten for more than a century, until President Cristina Kirchner conferred on her the title of General of the Argentinian Army in 2009, and in that same year, the Bolivian Senate promoted Juana Azurduy posthumously to the rank of Marshal of the Republic, declaring her “Liberator of Bolivia.” Although the biography of Juana Azurduy assuredly places her as one of the most important women of Latin America, the history of her fight for freedom and equality has not received enough attention outside history classes and political events. Naming Tropidurus azurduyae we do not aim to merely reverence her as a historical personage and revolutionary soldier, but to genuinely honor her intelligence, courage, and heroic actions against a male-dominated colonialist world whose roots remain alive at the present time. This is an affirmative action to remind all Latin American women and men of our female heritage of strength and combativeness.
For a more comprehensive biography of Juana Azurduy, refer to the work of the Argentinian writer Mario “Pacho” O’Donnnel (1994), available online (http://www.portaldesalta.gov.ar/ juana%20azurduy.htm). Those interested may follow the YouTube link (https://youtu.be/SERg- 8GKCNeA) to hear the song “Juana Azurduy” in the voice of the Argentinian singer Mercedes Sosa, honoring the valiant spirit of Juana Azurduy.
DISTRIBUTION, ENDEMISM, NATURAL HISTORY, AND CONSERVATION: Tropidurus azurduyae was discovered from its type locality, Torotoro National Park, Department of Potosí, Bolivia (figs. 1–2). There, it is abundant in the xerophytic inter-Andean valleys, not being found in adjacent habitats such as the prepuna, located at altitudes above ~ 2800 m. Specimens deposited at the Museo de Historia Natural Noel Kempff Mercado (Santa Cruz) and Museo de Historia Natural Alcide d’Orbigny (Cochabamba) revealed the occurrence of T. azurduyae in several other localities to the north, south, and east of Torotoro. Currently, the species is known from the Bolivian departments of Chuquisaca, Cochabamba, Potosí, and Santa Cruz, ranging from approximately 1040 to 2764 m. (fig. 2).
Tropidurus azurduyae is endemic to the Bolivian Montane Dry Forests ( Olson et al., 2001), restricted to inter-Andean dry valleys from central and southern Bolivia ( López, 2003a, 2003b; fig. 2). This is the first species of the T. torquatus species group (per Frost et al., 2001) endemic to the Andes, and it reaches the highest altitudes among all Tropidurus . Tropidurus melanopleurus Boulenger, 1902 , member of the T. spinulosus species group (per Frost et al., 2001), is the only other species in the genus recognized as an Andean endemic. However, it is found in more mesic habitats along the eastern Andean slopes, from river margins and foothills of 400–500 m to altitudes near 2000 m ( Laurent, 1982; Schumacher and Barts, 2003). Tropidurus melanopleurus ranges from southeastern Peru to northwestern Argentina, crossing the Bolivian territory from north to south predominantly along the Yungas and the Tucuman-Bolivian forests ( Meier, 1982; Cei, 1993; Dirksen and De La Riva, 1999; Rivadeneira, 2008; Carvalho, 2013). The species is known to be sympatric (but never syntopic) with T. azurduyae in just a few areas of inter-Andean dry valleys in the Department of Santa Cruz (e.g., La Angostura; E. Cortez, personal commun.). However, as in most of its distribution, it is restricted to quebradas (river margins), and thus it is absent in the harsh dry environments dominated by T. azurduyae .
The type locality and a few dry valley sites visited in Cochabamba are the only areas for which information on the natural history of T. azurduyae is available currently. The new species is heliophilous and basks over small to large rock blocks (40 cm to> 150 cm in diameter), either isolated or forming large rock aggregations throughout the dry valleys. Tropidurus azurduyae occasionally uses the trunk of the contorted trees that dominate the landscape. The dry valleys of Torotoro have reddish, stony soil similar in color to the flanks and gular region of the new species. Tropidurus azurduyae is territorial and reacts to invasion of its home range with aggressive head movements and body push-ups, but rapidly flees to holes underneath boulders, hides in crevices between rock blocks, or climbs up tree trunks if truly threatened. Our short visit to the type locality did not allow us to determine the exact period and pattern of activity of T. azurduyae ; however, lizards were seen active even during the hottest periods of the day. Specimens were collected in Torotoro from the second half of the morning (around 11 AM) to the second half of the afternoon (around 4 PM). In dry valleys sites from Cochabamba, T. azurduyae has been observed active throughout the whole year (including the winter), basking over rocks after approximately 8 AM. Nothing is known about the diet of T. azurduyae , but a few lizards were observed feeding on ants at the type locality. Most specimens collected by us had their mite pockets filled with a large number of bright orange chigger-mite larvae (fig. 3F), likely trombiculids, but the specific identity of these ectoparasites has not been investigated.
The conservation status of the Bolivian Montane Dry Forests has been defined as critical ( WWF, 2017), with habitat loss one of the most severe threats to this region ( Ibisch and Mérida, 2003; Aguirre et al., 2009; Navarro, 2011) and other seasonally dry tropical forests around the globe ( Janzen, 1988; Miles et al., 2006). Records of T. azurduyae (and other endemic taxa) obtained from museum specimens collected years or decades ago are, therefore, no guarantee that previously sampled populations persist to date. Although the local abundance of T. azurduyae in the protected Torotoro National Park and its broader distribution in the inter-Andean dry valleys from central and southern Bolivia indicate that the species is unlikely to be threatened, the lack of information about the size, connectivity, genetic parameters, and ecological requirements of local populations compels us to recommend its classification as “data deficient,” following the rules proposed by IUCN (2001).


| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Tropidurus azurduyae
| Carvalho, André L. G., Rivas, Luis Rolando, Céspedes, Ricardo & Rodrigues, Miguel T. 2018 |
Tropidurus azurduyae
| Carvalho & Rivas & Céspedes & Rodrigues 2018 |
T. azurduyae
| Carvalho & Rivas & Céspedes & Rodrigues 2018 |
T. azurduyae
| Carvalho & Rivas & Céspedes & Rodrigues 2018 |
T. azurduyae
| Carvalho & Rivas & Céspedes & Rodrigues 2018 |
Tropidurus azurduyae
| Carvalho & Rivas & Céspedes & Rodrigues 2018 |
Tropidurus azurduyae
| Carvalho & Rivas & Céspedes & Rodrigues 2018 |
T. azurduyae
| Carvalho & Rivas & Céspedes & Rodrigues 2018 |
T. azurduyae
| Carvalho & Rivas & Céspedes & Rodrigues 2018 |
T. azurduyae
| Carvalho & Rivas & Céspedes & Rodrigues 2018 |
Tropidurus melanopleurus
| Boulenger 1902 |
Tropidurus melanopleurus
| Boulenger 1902 |