
Tricrania sanguinipennis ( Say) 1824
|
publication ID |
https://doi.org/ 10.5281/zenodo.200535 |
|
DOI |
https://doi.org/10.5281/zenodo.6194902 |
|
persistent identifier |
https://treatment.plazi.org/id/FE334E68-B67E-A324-FF78-FE1FB6718B5D |
|
treatment provided by |
Plazi |
|
scientific name |
Tricrania sanguinipennis ( Say) 1824 |
| status |
|
Tricrania sanguinipennis ( Say) 1824 : 279
Horia sanguinipennis Say 1824: 279
Tricrania sanguinipennis: LeConte 1860: 320 ; Henshaw 1885: 130; Wellman 1910a: 219; Blatchley 1910: 1354; Borchmann 1917: 172; Leng 1920: 160; Blackwelder 1939: 35; MacSwain 1956: 136; Campbell 1991: 265
Type material. Say’s unique type specimen was presumably destroyed with his collection, and could not be located after an exhaustive search of museums and institutions. The authors expected the type to be located at the American Museum of Natural History, the Smithsonian Institution, or the Academy of Natural Sciences in Philadelphia. A neotype has not been designated as there has not been an exceptional need to do so (see Article 75.3 of the ICZN). The original type locality is Pennsylvania.
Adult redescription. Length: 8.5–15mm (mean = 11.5mm, N=20), width 3.5–5.5mm (mean = 4mm, N=20). Overall, body robust, nearly parallel-sided, convex, dark black to piceous with elytra brick-red, but elytra fading to a dull orange after death. Body surface covered with short black setae, most setae decumbent but sometimes erect on elytra, elytra more sparsely covered with setae than rest of body.
Head broadly triangular (W:L = 1.3:1); surface deeply densely punctate with irregular shaped punctures, some punctures coalescing. Punctures 1–2.5 diameter of eye facet; inner surface of punctures finely microreticulate; each puncture gives rise medially to an elongate fine decumbent seta; interspaces smooth to slightly granular, shining, <0.25 puncture diameters apart, more widely separated on temples. Antenna elongate, antennomere comparative lengths as follows: 1:0.66:1:1:1:0.85:0.85:0.85:0.66:0.66:1.33. Terminal antennomere longest, subequal to antennomeres 9 and 10 combined, apex acuminate. Antennomeres 1 and 3 widest, antennomere 2 distinctly shorter and more narrow than 1 or 3.
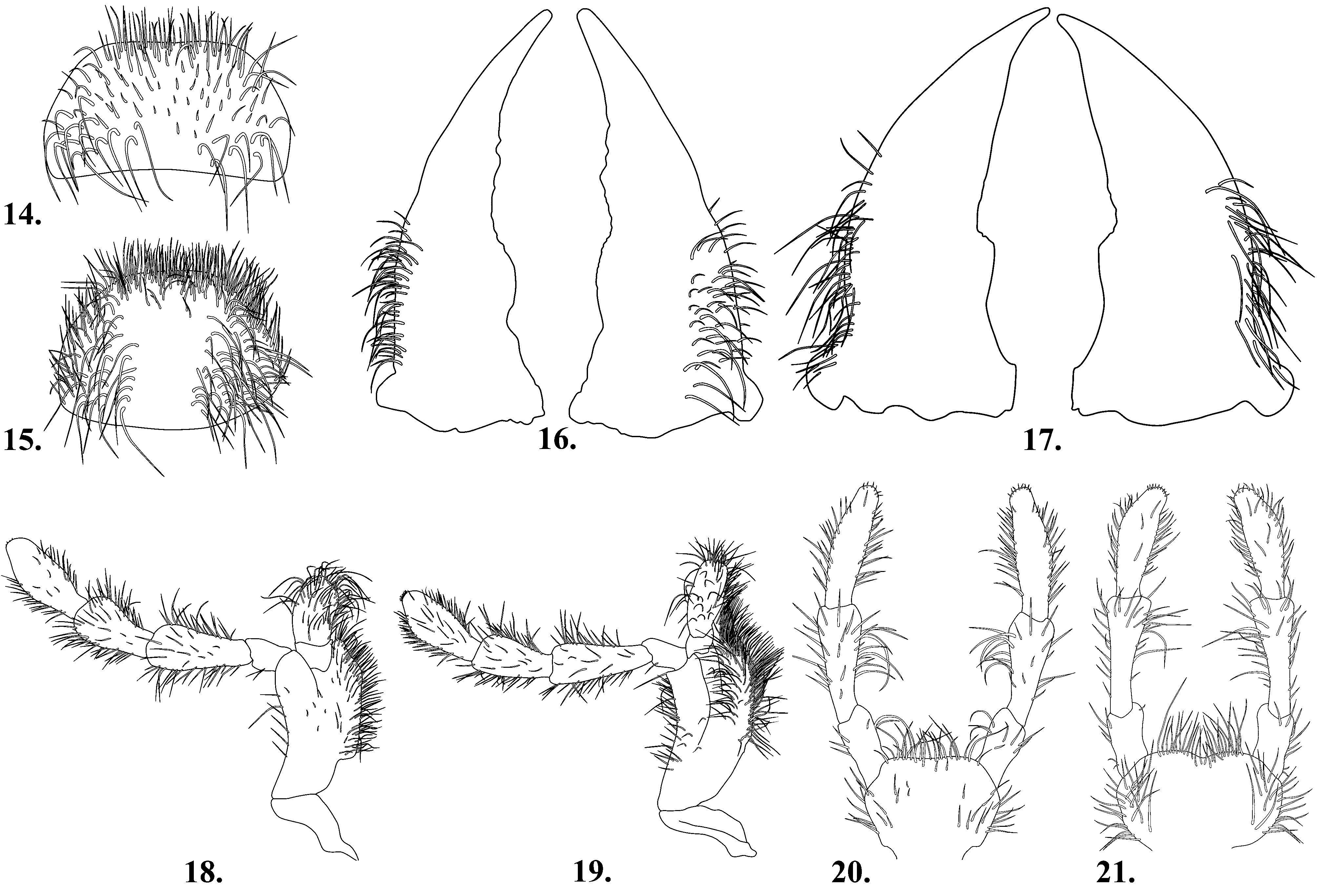
Mouthparts clearly visible, well developed. Labrum moderately transverse to subquadrate ( Fig. 14 View FIGURES 14 – 21 ); anterior margin appearing truncate, with moderately dense setal brush; posterior margin broadly concave. Mandibles large, somewhat slender, much longer than wide; lateral region with numerous projecting setae, more densely setose on right mandible; inner margin without well defined subapical tooth ( Fig. 16 View FIGURES 14 – 21 ). Maxilla with terminal palpomere bearing few scattered setae at apex; lacinia with lateral margin straight; galea with apex rounded ( Fig. 18 View FIGURES 14 – 21 ). Labium with anterior margin appearing truncate; labial palpus elongate, terminal palpomere longest, apex with few scattered sort setae, moderately swollen mesally ( Fig. 20 View FIGURES 14 – 21 ).
Pronotum moderately transverse, appearing subquadrate, widest near anterior angles (W:L = 1.3:1); lateral margins somewhat constricted in posterior 0.33; anterior and posterior angles broadly rounded; anterior and posterior margin broadly evenly convex. Surface punctures similar to those on head; interspaces distinctly finely microreticulate, shining. Scutellum with apex rounded; deeply densely punctuate as on head and pronotum; interespaces finely granular, moderately shining. Elytron with humeri poorly developed; sutural striae not apparent along entire length; apices moderately to narrowly rounded. Surface punctures indistinct, appearing wrinkled with alutaceous to granular sculpturing. Prosternum deeply densely punctuate, punctures small, half the size of those on head or pronotum; interspaces smooth to granular, shining. Mesosternum with distinct medial glabrous region in anterior 0.5 of structure, surface granular; punctures on remaining areas similar to those on prosternum. Metasternum similarly punctuate as pro- and mesosternum in posterior 0.5, more similar to head and protnoum in anterior 0.5 with larger punctures and interspaces more shining.
Legs elongate, well developed. Pro- and mesotibia with two unequal apical spurs, anterior spur straight and posterior spur apically curved. Metatibial spurs unequal, both appearing flattened and spoon shaped with the anterior spur more so. Tarsal claws complex, each claw with a ventral blade subequal to length of dorsal blade, each dorsal blade also finely denticulate from apex to near base.
Ventrites 1–5 large, subequal in length to each other; punctures large and similar to those on head and pronotum, becoming more densely spaced laterally; interspaces distinctly microreticulate and granular, moderately shining.
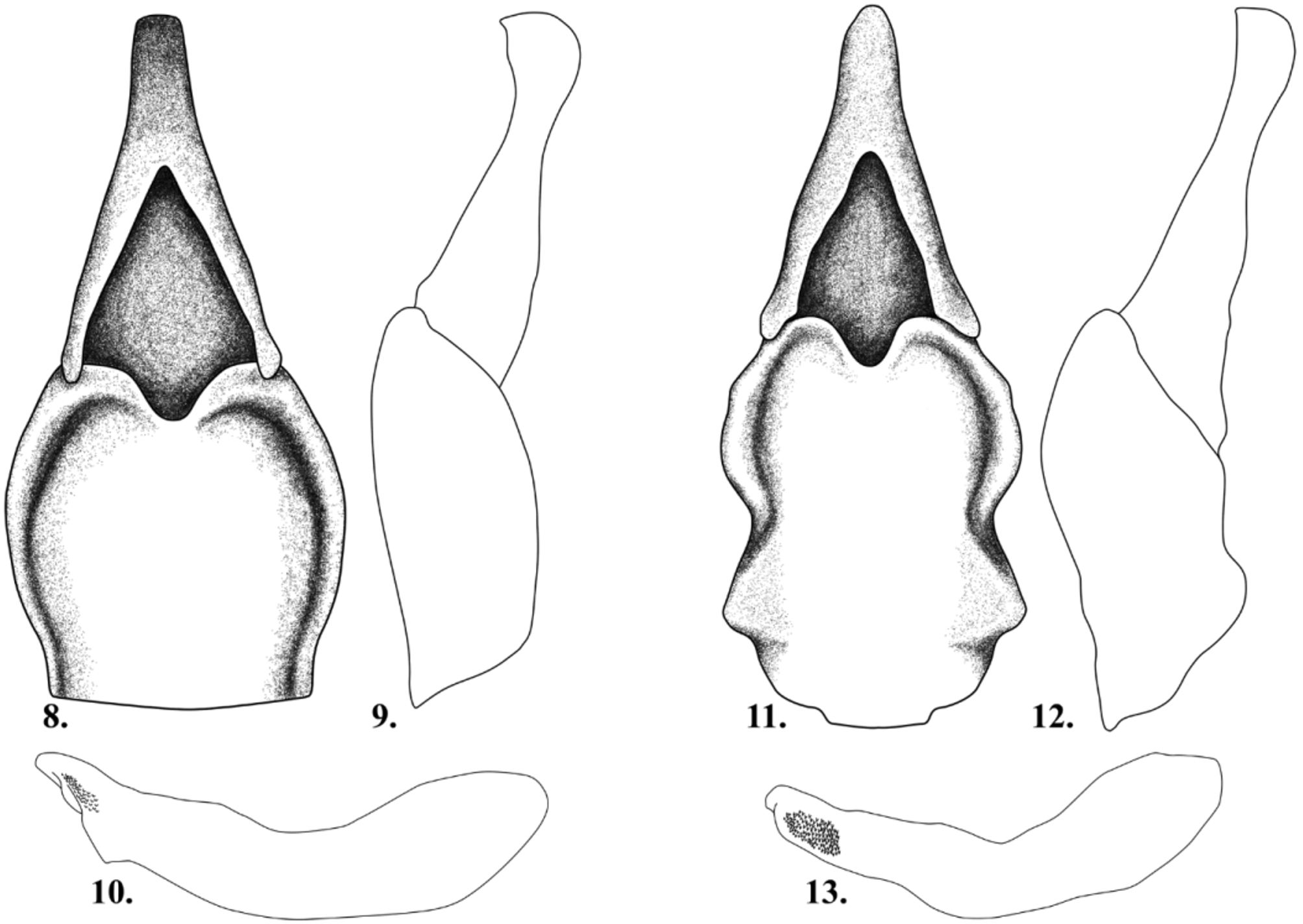
Male genitalia well sclerotized. Tegmen with parameres fused, and articulated with phallobase ( Figs. 8 View FIGURES 8 – 13 ). In ventral view, parameres bluntly rounded apically, with deep medial fossa and deeply convex attachment to phallobase; phallobase wider than parameres, widest near middle, median fossa broadly shallow. In lateral view, parameres with sharp apical point and phallobase evenly convex ( Fig. 9 View FIGURES 8 – 13 ). Median lobe of aedeagus in lateral view with moderate declivity from apex to base, sharply defined tooth-like dorsal projection, and small apical field of minute projections ( Fig. 10 View FIGURES 8 – 13 )
Female genitalia weakly to moderately sclerotized. Species level diagnostic characters unapparent for genitalia.
Variation. Little variation observed in color, punctation and relative shape and size of male and female genitalia; however, notable differences were observed in overall length, width, and convexity among individuals, as well as the degree to which the pronotum is transverse with some individuals exhibiting a W:L = 1.45:1, the number of denticles on each tarsal claw also varies. Females were generally larger than males.
Etymology. The specific epithet is a combination of the Latin “sangui” meaning blood, and “penni", meaning wing; apparently this epithet is indicative of the dark red elytra of the species.
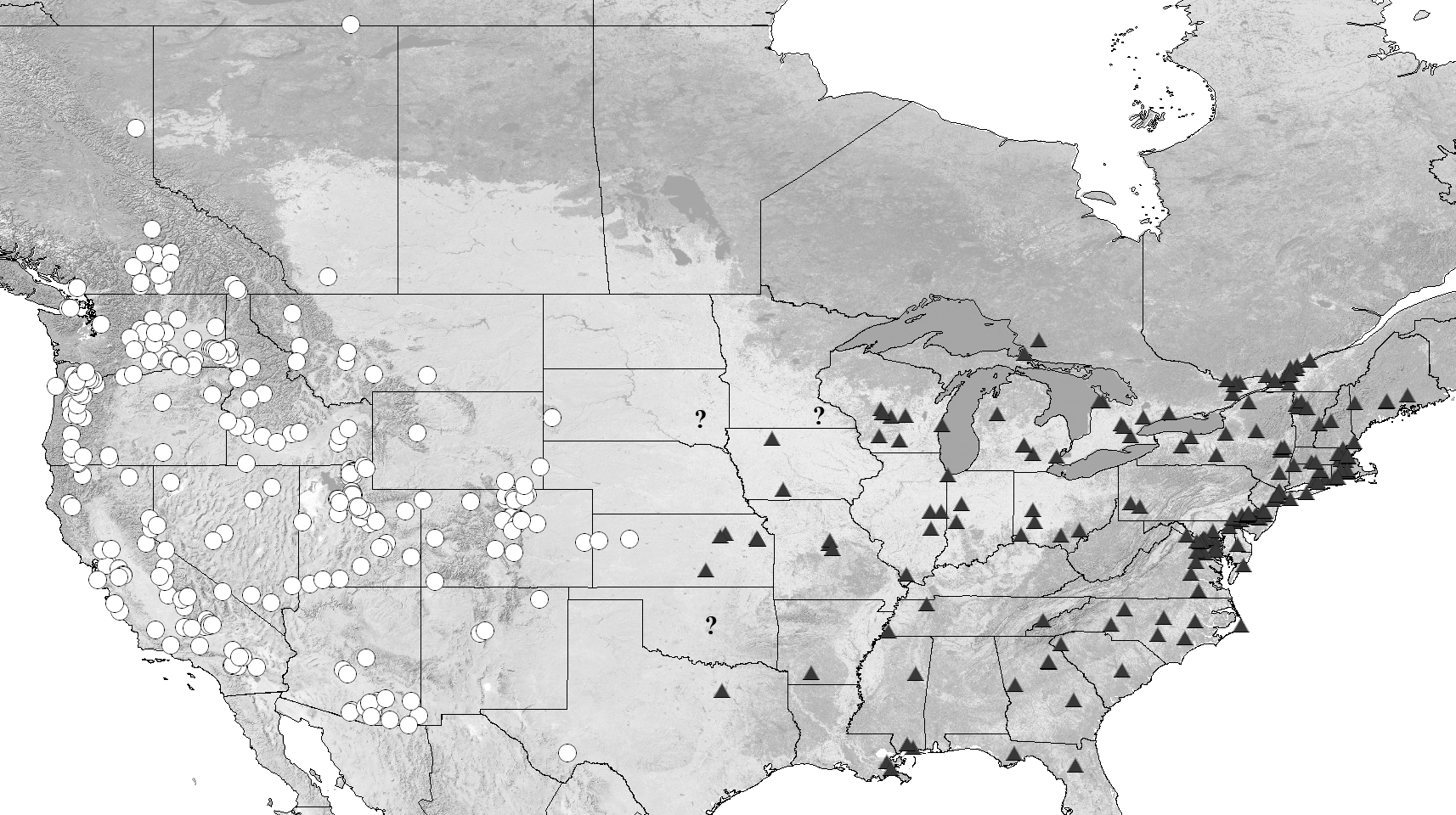
Distribution. Eastern North America ( Fig. 30 View FIGURE 30 ), including: Canada (QC, ON) and the United States (AR, CT, DC, DE, FL, GA, IA, IL, IN, KS, LA, MA, MD, ME, MI, MO, MN, MS, NC, NH, NJ, NY, OH, OK, PA, RI, SC, SD, TN, TX, VA, VT and WI). One record from BMNH is listed as “Mex”; however, as mentioned above, the record is somewhat dubious. An exhaustive search at Mexican institutions, and persons and institutions known to have made collecting trips to Mexico, yielded no additional Mexican records. We believe populations may exist in Mexico, however more fieldwork is required to test this. Therefore, Mexico is not indicated in the distribution map.
Question marks for South Dakota and Oklahoma represent specimens labeled with only a state name, and for which no further records exist. The question mark in Minnesota represents a locality record from Olmsted County given in an unpublished thesis ( Welles 1968), and for which no voucher could be located.
Biology. The first robust treatment of this species was provided by Parker and Böving (1924); however, we have greatly expanded on these initial observations (see Table 2 below). A slide of triungulins from MacSwain’s collection at the University of California - Berkeley also provides some insight into the biology of this species. The slide noted that adults were collected “in copula” in mid April, eggs were deposited six days later, and subsequent first instar larvae hatched approximately three weeks after oviposition in mid May.
For several years, one of us (JPH) has recorded detailed biological information for T. sanguinipennis at a sand pit located in the southern part of the New York Adirondack State Park. The pit is ~ 3 miles east of the junction of Routes 8 and 365 in Herkimer Co., NY. The floral composition of the area is considered southern Adirondack forest. The sand pit itself probably has not been mined for 20–30 years. The microhabitat of the pit includes coarse sand with small smooth pebbles intermixed with occasional small smooth rock (potential glacial outwash). The south facing north end has clumped small (2–3 meter) hardwood trees and bushes with dispersed moss cover. In these areas, solitary ground bee burrows have been numerous, but much less common elsewhere, especially on both the east and west flanks of the pit. Small pieces of boards and other human-based debris along with sticks and small logs are scattered on the south facing north slope.
One observation on April 21, 2005 yielded a single dead specimen under a small piece of board. However, the following year on April 11 and 20, 56 specimens were found in the same place. On both days the weather was cool (~15°C) with clear skies and the sun intensely shining on the slope face. Collection events occurred between 11:00–1:00 pm on both days. Ground bees were numerous and extremely active, even though some snow was present in the surrounding woods. Of the 56 beetles collected, 63% were found live under small board pieces and/or sticks, 20% dead, 10% dead and lightly webbed by spiders, 6% dead on the sand, and 1% walking on the sand. All live and spider-webbed individuals under wood were upside down under the board.
Return trips on May 1 and 3, 2007 yielded 298 specimens. Visits in May rather than April were due to late snowfall and an unusually cold late spring in the area. Both stops were only an hour in length, at around 11:00am. A thorough search of the pit on both dates for more specimens easily would have yielded 100–200 additional beetles. Of the 298 individuals, 148 were found dead on the sand or in small depressions, 128 were found live under wood, 32 were found dead under wood (killed by spiders, i.e. cocooned), and 17 were found crawling on the sand. Most live individuals under the boards, sticks, and small logs were producing large egg masses. Samples of live individuals were collected for molecular vouchers. Two spiders apparently feeding on the beetles also were collected as well as host ground bees ( Colletes inequalis Say ). Numerous other spiders of the same species were discovered under the pieces of wood. Bees were quite active due to the warm weather with partly cloudy to clear skies. There appeared to be a dramatic increase of solitary bee burrows from 2005 to 2007, but the nests were more widely separated than in previous years at the prime collecting areas. No copulating pairs of beetles were observed from 2005 or 2007, thus it remains unclear when mating occurs. The method for the triungulin larvae to locate and invade ground bee borrows several inches to feet from the egg mass sites remains unknown, but presumably involves phoresy.
Another set of observation events occurred on April 17 and 25, 2008, at which time there were already freshly dead individuals on the sand, others on their back, kicking but near death, a few out walking rapidly on the surface, and a number under pieces of wood, but with no evidence of eggs. Five pairs were found together, but not in copulation at that time, e.g. 12:00–1:00 pm. No "pairs" were found of the hundreds observed in 2007. In 2008 the spiders were apparently killing and wrapping up only pairs of Tricrania , which we cannot explain. On 25 April the Colletes were estimated at about 40% of the 2007 population level, and Tricrania about 20% of the 2007 population. The dramatic drop in Colletes population levels was not surprising given the extreme pressure put upon them by the immense Tricrania boom in 2007. Five live spiders were collected, which had lightly webbed (very few silken strands) Tricrania individuals for eventual feeding. The spiders were kept alive to allow them to mature. Two Tricrania were still laying eggs in the typical manner (i.e. upside down on a wooden surface). The adult Colletes , in the early morning as they first crawl from the burrows, actively run about on the ground near the burrow entrance. However, during the day active flight is most typical with moderately frequent landings on the sand (short durations of a few seconds) and then the bees are airborne again. On the 25 April, as the Colletes were walking around their burrows, the air temperature was ~ 10˚C, but the ground likely was much cooler, except where direct sunlight was shining on the soil surface. That day was cloudless between 9:00a.m. and 10:00a.m. and the temperature near the sand areas receiving full sunlight may be much warmer than the surrounding shaded areas by as much as -7˚C.
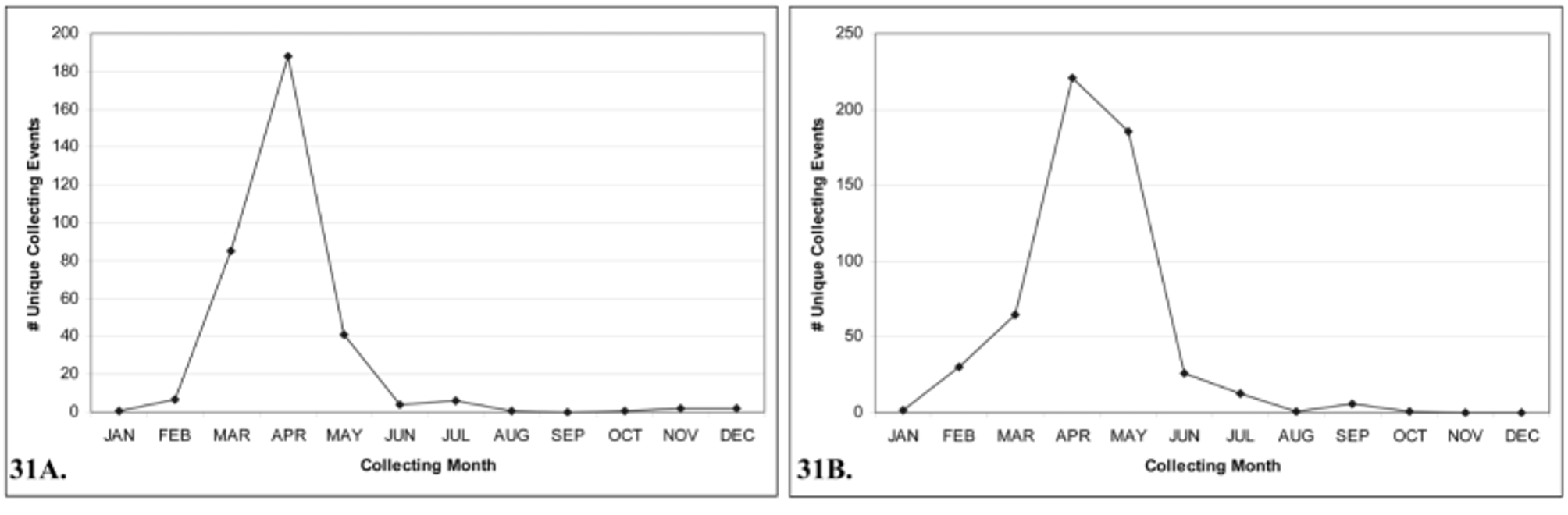
According to our data, there is a definitive seasonality associated with both Tricrania species ( Figs. 31 View FIGURE 31 A, 31B). Both species have short-lived spring active adults, which likely coincides with initial activity of ground dwelling bees and therefore soil temperature. Data from>300 unique collecting events for T. sanguinipennis suggest a peak in adult activity from March through early May; beginning near mid-March and extending into early-May.
Notes. This species was originally described from a single specimen by Say (1824), who remarked that, “…it must be rare, only a single specimen having yet occurred”. Although rare in collections, we note here that this species is locally abundant, being collected in great abundance in sandy locations, but is temporally ephemeral in occurrence. Enns (1956) inferred that this species was restricted to the single Hymenoptera genus Colletes ; however as noted in Table 1, triungulins of this species are phoretic on at least three bee genera. The first instar larval description and diagnosis has been previously completed in detail ( MacSwain 1956, Parker & Boving 1924), and will not be repeated herein; however, illustrations are provided ( Figs. 22, 24, 26 View FIGURES 22 – 29 , & 28). We have included molecular sequence data, i.e. COI barcode region, in Appendix 4 as an initial step to promote further explorations on the biology, systematics, and evolution of this species.
TABLE 1. Faunal, floral and miscellaneous collecting records for T. sanguinipennis (Say) * 1 Parker & Böving 1924, Erickson et al. 1976; 2 Stephen 1954, Erickson et al. 1976; 3authors pers. obs. or label data from Appendix 2 below. *These records do not necessarily reflect rearing records.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Tricrania sanguinipennis ( Say) 1824
| Cline, Andrew R. & Huether, Jeffrey P. 2011 |
Tricrania sanguinipennis:
| Campbell 1991: 265 |
| MacSwain 1956: 136 |
| Blackwelder 1939: 35 |
| Leng 1920: 160 |
| Borchmann 1917: 172 |
| Wellman 1910: 219 |
| Blatchley 1910: 1354 |
| Henshaw 1885: 130 |
| LeConte 1860: 320 |
Tricrania sanguinipennis (
| Say 1824: 279 |
Horia sanguinipennis
| Say 1824: 279 |