
Triaenops pauliani, Goodman & Ranivo, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.5755724 |
|
persistent identifier |
https://treatment.plazi.org/id/085A87FC-BE1D-FFB6-FF3D-FCE2CB2088D4 |
|
treatment provided by |
Marcus |
|
scientific name |
Triaenops pauliani |
| status |
sp. nov. |
Triaenops pauliani View in CoL n. sp.
( Figs 2 View FIG ; 3 View FIG ; Tables 1; 2)
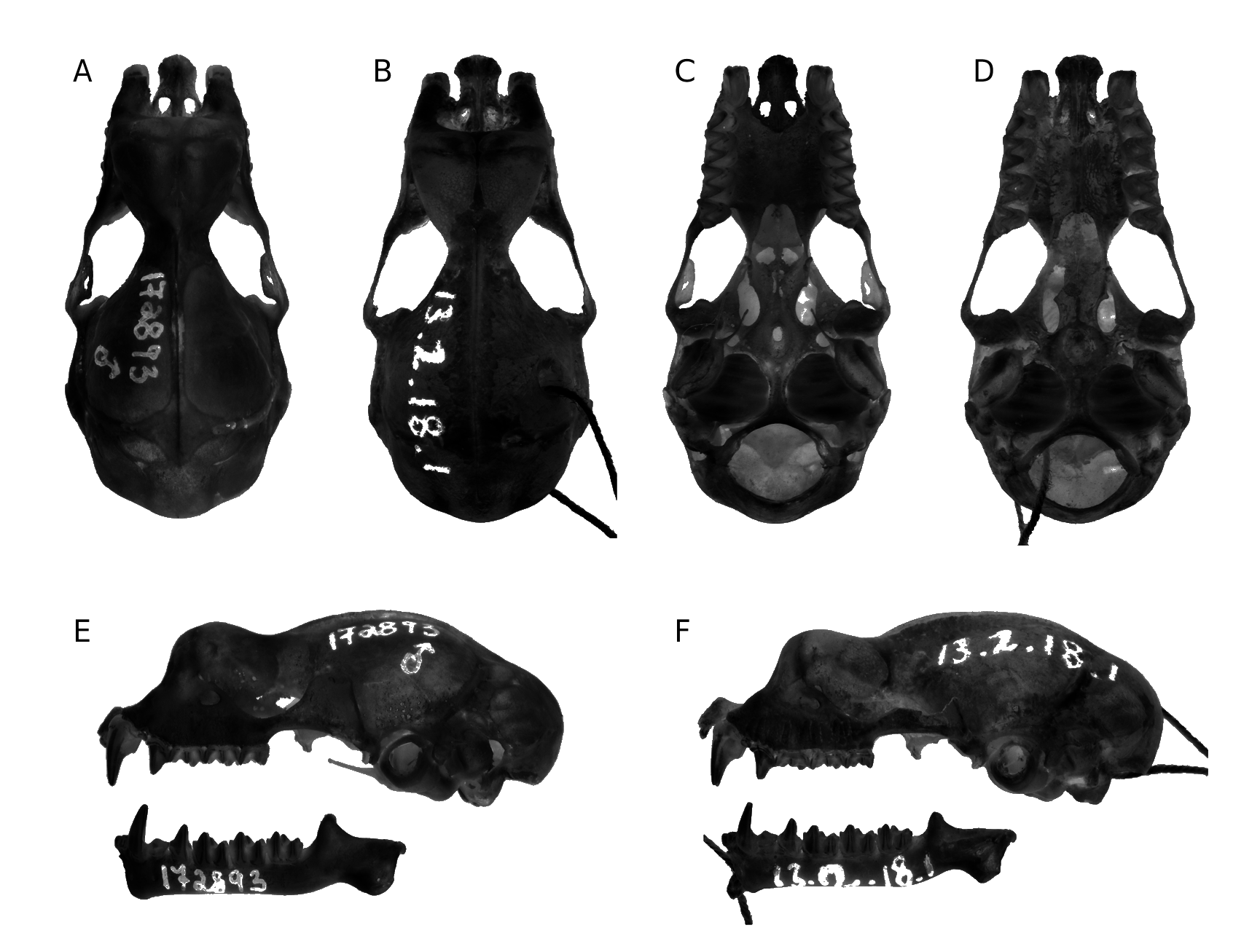
HOLOTYPE. — Adult ♂, date not specified, J. C. F. Fryer coll. ( BMNH 13.2.18.1). Based on the itinerary of the Percy Sladen Trust Expedition to the Seychelles, Fryer was in the western Seychelles (Astove, Cosmoledo, Assumption, and Aldabra) from 13.IX.1908 to 24.I.1909 ( Gardiner et al. 1910).
The specimen was presumably preserved in formalin and subsequently transferred into alcohol. The skull was extracted and cleaned. The cadaver is generally in a good state of preservation, as is the skull, with the exception of the right pterygoid plate that has been damaged and largely broken. A hole has been drilled into the back of the cranium for the insertion of a thread used to attach a specimen tag and the mandible.
PARATYPES. — Aldabra Atoll , Picard Island, c. 09°24’S, 46°12’E, 13.IX.1908 - 24.I.1909, J. C. F. Fryer coll., fluid preserved specimen with skull removed, dorsal portion of cranium missing, upper toothrows complete, complete mandible, 1 adult ♀ ( BMNH 13.2.18.2). — Same data, fluid preserved specimen, 1 adult ♀ ( CUMZ E5609 View Materials .A) GoogleMaps ; 1 adult ♂ ( CUMZ E5609 View Materials .B). — Aldabra Atoll , Picard Island , Aldabra Station , 09°24’04.4”S, 46°12’21.4”E, 5 m, 4.V.1977, J. J. Whitelaw coll., fluid preserved specimen with skull in situ, 1 adult ♀ ( BMNH 78.185 ). — Aldabra Atoll, Picard Island, Aldabra Station, 09°24’04.4”S, 46°12’21.4”E, 9.IV.2004, U. Samedi coll., fluid preserved specimen with skull removed, 1 sub-adult ♂ ( FMNH 185795 About FMNH ) GoogleMaps .
ETYMOLOGY. — The name pauliani is a patronym after the late Prof. Renaud Paulian, who conducted extensive research in the western Indian Ocean islands and was responsible for a major synthesis of the biogeography of Madagascar and neighbouring islands ( Paulian 1961). Previously Tate (1941: 3) used the name “ Triaenops furinea Trouessart ” for the species on Aldabra. To our knowledge Trouessart never named the form occurring on Aldabra; the use of “ furinea ” is presumably a misreading of the name furcula and is considered a nomen nudum ( Hayman & Hill 1971: 30; Hill 1982).
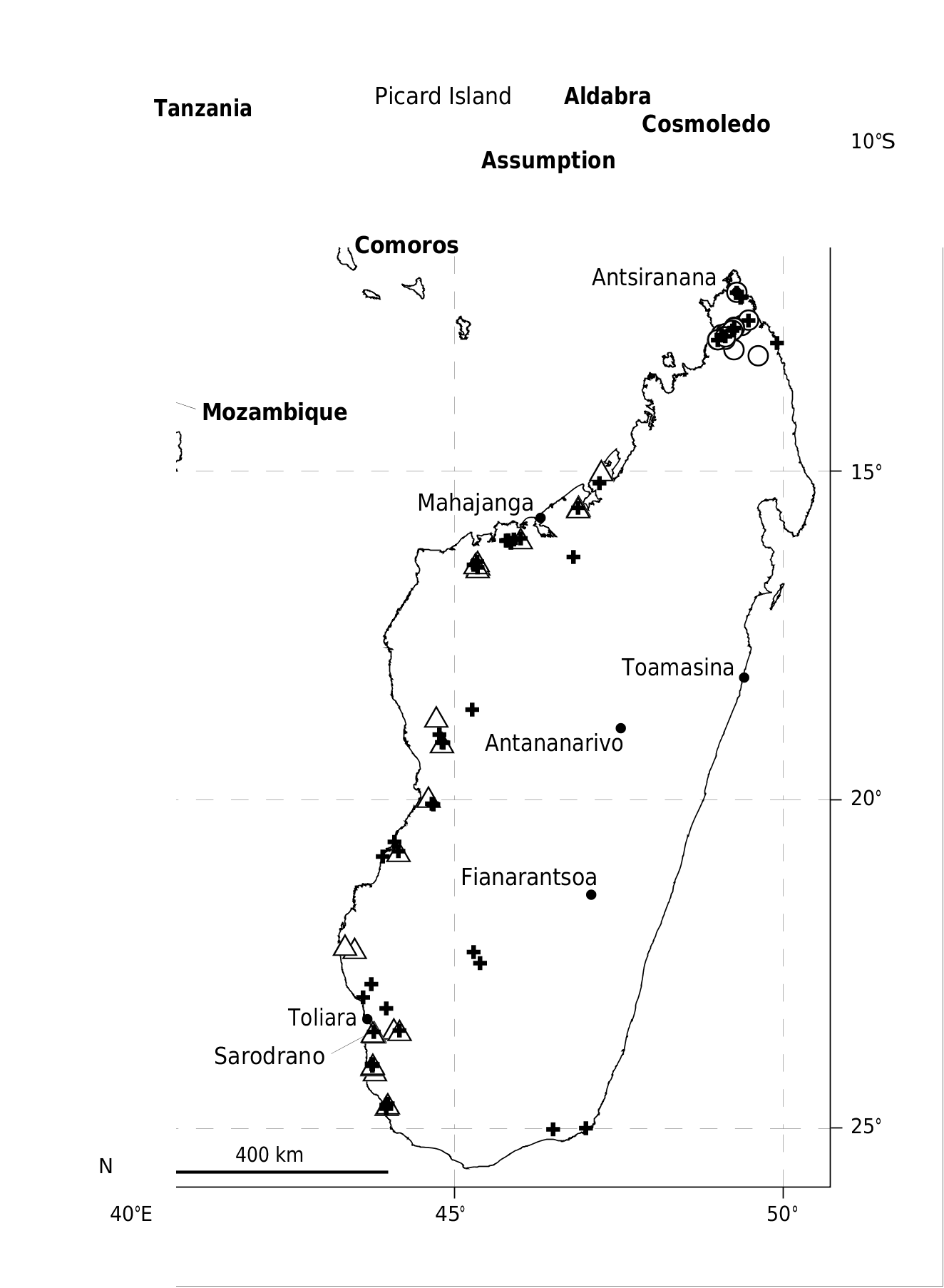
TYPE LOCALITY. — Aldabra Atoll, Picard Island, c. 09°24’S, 46°12’E ( Fig. 1 View FIG ) (see Discussion for precision of holotype locality).
DISTRIBUTION. — Triaenops pauliani n. sp. is currently known only from the western portion of the Seychelles Archipelago, specifically the Aldabra Atoll, Picard Island. Previous records of this taxon on the Cosmoledo Atoll are called into question (see Discussion). The nearest landmass to the Aldabra Atoll is Assumption Island, 27 km to the south.
HABITAT. — No precise information is available on the site where the holotype and several of the paratypes were collected. Picard Island, the only known site where this species is clearly documented from, is in the northwestern portion of the Aldabra Atoll, which is divided into four different islands. The Aldabra Research Station is located on Picard Island, has been the focal point of faunistic studies on the atoll, and is in close proximity to an abandoned settlement. One of the specimens (FMNH 185795) was obtained in a house within the research station complex, where it was presumably attracted after sunset to insects flying around an electric light (Samedi pers. comm.). The islands making up the Aldabra Atoll complex are uplifted fringing coral reefs that rise a few meters above sea level and Stoddart et al. (1971) have described their geomorphology in detail. The climate is semi-arid with the rainy season falling between November and April and summer maximum temperatures average 32°C and winter minimum temperatures average 22°C – in general, the local climatic regime is similar to Antsiranana in northern Madagascar ( Farrow 1971).
The terrestrial vegetation of Picard Island is relatively intact, composed of several different formations ( Stoddart & Wright 1967): mixed scrub – a variable formation in the floristic sense and often with some introduced plants near the settlement; Pemphis thicket – dominated by P. acidula ; psammophilic communities – a dwarf plant community found close to the shore or at edge of cliffs; man-induced vegetation – principally coconut plantations, Casuarina thickets and woodlands, and other non-native vegetation.
MEASUREMENTS. — Measurements taken directly from the fluid preserved holotype.
External (approximate): total length 55 mm, tail length 18 mm, hindfoot length 5.8 mm, ear length 15.3 mm, forearm length 43.7 mm, external noseleaf length 3.8 mm, central noseleaf length 3.7 mm.
Skull and teeth: GSKL 17.1 mm, ZYGO 8.7 mm, IOB 2.3 mm, MAST 8.3 mm, PAL 4.4 mm, MAND 11.1 mm, I 1 -M 3 6.9 mm, MOLS 4.4 mm, C 1 -C 1 4.2 mm, and M 3 -M 3 6.1 mm ( Table 2).
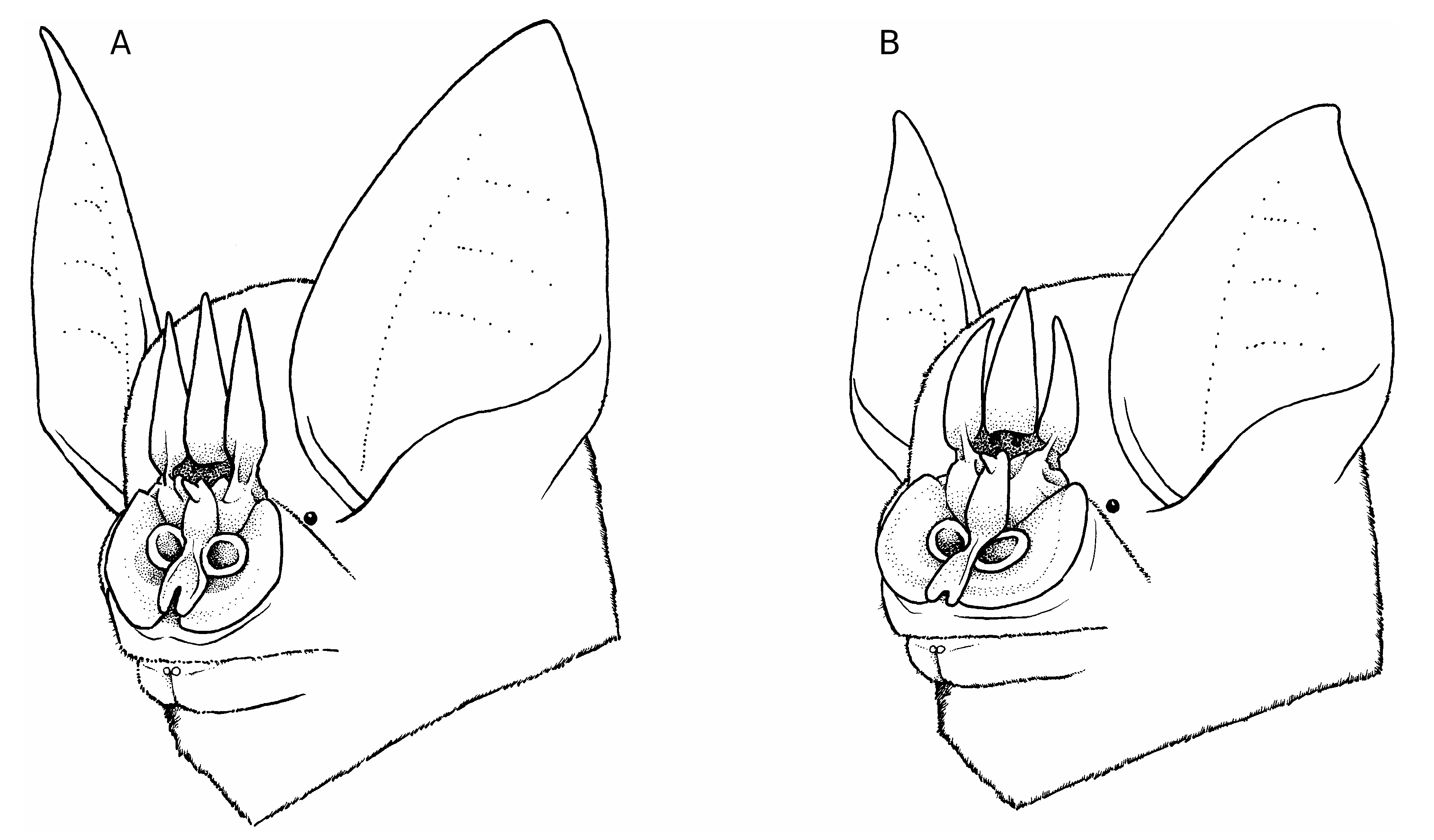
DIAGNOSIS. — Distinctive cranial and external characteristics, including the dental formula 1/2-1/1-2/2-3/3, inflated rostrum, wide plated zygoma, thickened premaxilla, and distinct trident noseleaf, clearly identify the Picard Island animals as members of the genus Triaenops ( Hill 1982; Koopman 1994). The noseleaf structure of the western Seychelles specimens, with three elongated tridents, nearly the same length, places this animal within the T. furculus / auritus species group, as compared to the T. rufus / persicus group where the outer noseleafs are shorter than the internal one and distinctly different ( Hill 1982). Triaenops pauliani n. sp. is distinguished from T. furculus and T. auritus on the basis of slightly concave and less elongated outer noseleaf lancets ( Fig. 2 View FIG ); diminutive general body size, including a distinctly shorter ear length (14.5-15.4 mm); and a less inflated rostral swelling ( Fig. 3 View FIG ).
DESCRIPTION
Within the Malagasy species of Triaenops , there are two distinct types of noseleafs. Specimens referable to T. rufus have the central lancet of the trident noseleaf as an elongated spear-like structure and the two outer lancets are distinctly shorter than the central lancet and notably concave ( Peterson et al. 1995: 83). The trident structure in T. persicus of Africa and the Middle East ( Hill 1982: 179), the only other previously recognized species in this genus living outside Madagascar ( Simmons 2005), is similar to T. rufus . These two animals are each other’s closest living relatives ( Russell et al. 2007). In contrast, individuals referable to T. auritus and T. furculus have a more elongated central lancet than in T. rufus and the two outer projections of the trident are distinctly lance-shaped and almost equal in length to the central lancet ( Peterson et al. 1995: 80). Based on these characters, T. pauliani n. sp. is a member of the Malagasy T. auritus / furculus group.
The noseleafs of T. pauliani n. sp. are shorter than those in individuals of T. auritus or T. furculus , with, for example, the central noseleaf in male T. furculus measuring on average 5.3 mm (n = 15), in male T. auritus 4.9 mm (n = 15), and in five adult individuals of T. pauliani n. sp. 3.4 mm (♀), 3.7 mm (♂), 3.7 mm (♀), 4.0 mm (♀), and 4.0 mm (♂). Further, in T. pauliani n. sp. the external lancets are shorter than the internal lancet, and the outer margins of the external lancets are distinctly arcshaped ( Fig. 2 View FIG ), while in members of the T. auritus / furculus group the external lancets are distinctly spear-shaped and the difference in length of the external and internal lancets is less marked. In the hand, these characters readily distinguish T. pauliani n. sp. from these other two species.
Finer details of the noseleaf structure of T. pauliani n. sp. are difficult to discern from BMNH 13.2.18.1 and 13.2.18.2, in part associated with the fact the skulls have been extracted and the considerable period since their original preservation rendering some of the structures flaccid. However, in BMNH 78.185 and CUMZ E5609.A and E5609.B the skull remains in each respective specimen and the horseshoe of the noseleaf is notably broader than in typical specimens of the T. auritus / furculus group and distinctly more rounded along the lateral margin ( Fig. 2 View FIG ). Further, the anteriormost portion of the secondary horseshoe in T. furculus is notably more indented than in T. pauliani n. sp.
Other external characteristics separate T. pauliani n. sp. from members of the T. auritus / furculus group. In the latter species complex, most external measurements are larger than in T. pauliani n. sp. including, for example, ear length ( Table 1), which on average in males of T. furculus is 18.7 mm (n = 6), in males of T. auritus is 17.9 mm (n = 17), and in five individuals of T. pauliani n. sp. is 14.5 mm (♀), 14.6 mm (♂), 14.9 mm (♀), 15.3 mm (♂), and 15.4 mm (♀). Further, the hindfoot lengths of T. pauliani n. sp. in the males are 5.7 mm and 5.8 mm and in three females 6.0 mm, 6.0 mm, and 6.2 mm, as compared to the average measurements in males and females (respectively) in T. furculus of 6.8 mm (n = 6) and 7.5 mm (n = 4) and in T. auritus of 7.4 mm (n = 16) and 7.7 mm (n = 36). The only datum on body mass of T. pauliani n. sp. is from the male specimen obtained by Whitelaw in 1977 (BMNH 78.185), which weighed 6.5 g ( Brandis 2004); this falls within the range of T. furculus from Madagascar ( Ranivo & Goodman 2006).
One of the distinguishing characters separating adult specimens of T. auritus and T. furculus is the pelage coloration. In adult specimens of the former species the fur is generally reddish-gold, while that of T. furculus is brownish-gray or occasionally light yellowish-gray; in both species the dorsum is darker and more saturated with colour than the ventrum ( Ranivo & Goodman 2006 and unpubl. data). The four specimens of T. pauliani n. sp. collected by Fryer (BMNH 13.2.18.1 and 13.2.18.2, CUMZ E5609.A and E5609.B) have been stored in alcohol for close to 90 years and it is presumed that the pelage pigmentation has changed dramatically from their original natural colour. In contrast, BMNH 78.185, an adult male, and FMNH 185795, a subadult male, were collected and preserved over the past 30 years and presumably display a more natural pelage coloration. In BMNH 78.185, the dorsum is brownish-gray and the ventrum mouse gray, typical of adult T.furculus and in FMNH 185795 the pelage is a light gray, typical of sub-adult T. furculus .
The cranial measurements of T. pauliani n. sp. are similar to those of T. auritus and T. furculus and fall within the range of size variation of these latter two taxa ( Table 2). The same point holds in general for different dental measurements, although there is an elongation of the anterior portion of the upper toothrows in T. pauliani n. sp., with the I1-M3 measurements (6.9 [♂], 7.0 [♀] mm) falling outside the range for T. furculus (6.0- 6.4 mm, sexes combined, n = 16) and T. auritus (6.1-6.8 mm, sexes combined, n = 53). In contrast, the molar toothrows are generally shorter in T. pauliani n. sp. (4.4 mm [♂], 4.4 mm [♀]) than in T. furculus (4.5-4.9 mm, sexes combined, n = 16) or T. auritus (4.5-5.2, sexes combined, n = 53).
The morphology of the crania of T. pauliani n. sp. and T. auritus and T. furculus are similar to one another ( Fig. 3 View FIG ). One consistent difference, based on the limited number of cranial specimens available of T. pauliani n. sp., is that in this taxon the rostral area is less inflated, particularly the narial compartments, than in individuals of T. auritus and T. furculus . When seen in side view, the inflated rostrum in T. pauliani n. sp. does not rise to the level of the braincase, while in T. auritus and T. furculus the swelling is at a level equal or surpassing the braincase. In both T. auritus and T. furculus the rostral depression is more prominent than in T. pauliani n. sp. Further, the sagittal crest terminates anterior to the interorbital constriction in T. pauliani n. sp., and generally posterior to the constriction in T. auritus and T. furculus .
| CUMZ |
Cameroon University, Museum of Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |