
Sarika siamensis (Pfeiffer, 1856)
|
publication ID |
https://doi.org/ 10.1163/18759866-BJA10027 |
|
DOI |
https://doi.org/10.5281/zenodo.8348975 |
|
persistent identifier |
https://treatment.plazi.org/id/038587FD-0366-FFC8-745B-FAE89782FED9 |
|
treatment provided by |
Felipe |
|
scientific name |
Sarika siamensis (Pfeiffer, 1856) |
| status |
|
Sarika siamensis (Pfeiffer, 1856) View in CoL
( figs 1 View FIGURE 1 , 2 View FIGURE 2 , 3C, D View FIGURE 3 , 4C, D View FIGURE 4 , 6 View FIGURE 6 , 7 View FIGURE 7 , 11B View FIGURE 11 )
Helix siamensis Pfeiffer, 1856a: 32 . Type locality: Siam [ Thailand] .
Helix (Nanina) mouhoti Pfeiffer, 1860: 136 , pl. 50, fig. 5 View FIGURE 5 . Type locality: Siam [ Thailand]. Pfeiffer 1868: 116, 117.
Helix siamensis: Pfeiffer, 1856b: 76 , 77, pl. 21, figs 7–9 View FIGURE 7 View FIGURE 8 View FIGURE 9 . Pfeiffer, 1859: 60; Morelet, 1875: 247, 248.
Nanina siamensis: Marten, 1860: 8 ; Martens, 1867: 71, pl. 6, fig. 6 View FIGURE 6 ; Tryon, 1886: 86, pl. 16, figs 37–39.
Nanina birmana: Marten, 1860: 7 (misidentified of Pfeiffer).
Nanina hainesi: Marten, 1860: 7 (misidentified of Pfeiffer).
Nanina mouhoti: Tryon, 1886: 85 View in CoL , pl. 16, fig. 32. Hemiplecta dichromatica Morlet, 1889: 124 View in CoL , 175, 176, pl. 6, fig. 2 View FIGURE 2 . Type locality: de Srakéo à AngSon ( Siam) [Srakeo Province, Thailand].
Nanina (Xesta) siamensis: Fischer & Dautzenberg, 1904: 394 .
Xestina siamensis: Tomlin, 1932: 319 , 320, pl. 27. Nanina (Hemiplecta) siamensis: Kobelt, 1905: 1136 , pl. 285, figs 10 View FIGURE 10 , 11 View FIGURE 11 .
Hemiplecta siamensis: Solem 1966: 27 ; Panha, 1996: 34; Hemmen & Hemmen, 2001: 44.
Naninia [sic] mouhoti: Hemmen & Hemmen, 2001: 45 .
Cryptozona siamensis: Panha, 1996: 34 View in CoL ; Hemmen & Hemmen, 2001: 43; Prasankok & Panha, 2011; Abu-Bakar et al., 2014; Tan et al., 2016; Inkhavilay et al., 2019: 75 View Cited Treatment , 76, figs 35a, 56b; Boonmachai & Nantarat, 2020; Dumidae et al., 2020; Sutcharit et al., 2020b: 24 View Cited Treatment , 25, fig. 4d View FIGURE 4 ; Pholyotha et al., 2021.
Material examined. Type material: the type specimens could not be traced. Non-type material: see supplementary table S5.
Diagnostic characteristics. Shell depressed; shell colour generally two-tones (brownish above) or monochrome whitish to brownish; shell ribbed with regular-sized, dash-shaped nodules arranged on radial ribs. Animal with four mantle edges. Genitalia with straight epiphallic caecum and cuboidal penial pilasters. Spermatophore: head filament with smooth longitudinal folds; tail filament bearing three spines near sperm sac and about seven-tenths of its length with branching spines. Description. Shell ( fig. 3C, D View FIGURE 3 ). Shell depressed to conoid-depressed, large (width
up to 38.5 mm, height up to 20.5 mm), and moderately solid. Shell colour generally two-tone with yellowish to dark brown above and whitish below periphery (seldomly monochrome whitish to brownish), and brownish peripheral band sometimes present. Whorls 6–6½, increasing normally. Spire dome-shaped; apex slightly protruding; suture impressed. Embryonic shell nearly smooth with very weak radial wrinkles. Last whorl large and well-rounded. Upper shell surface sculptured with regular and distinct radial ribs, crossed by spiral lines and with series of regular-sized dash-shaped nodules arranged on radial ribs, and then radial ribs gradually disappearing below periphery towards umbilicus. Aperture crescent-shaped, obliquely opened with simple peristome. Columellar margin simple and slightly reflected near umbilicus. Umbilicus narrowly opened.
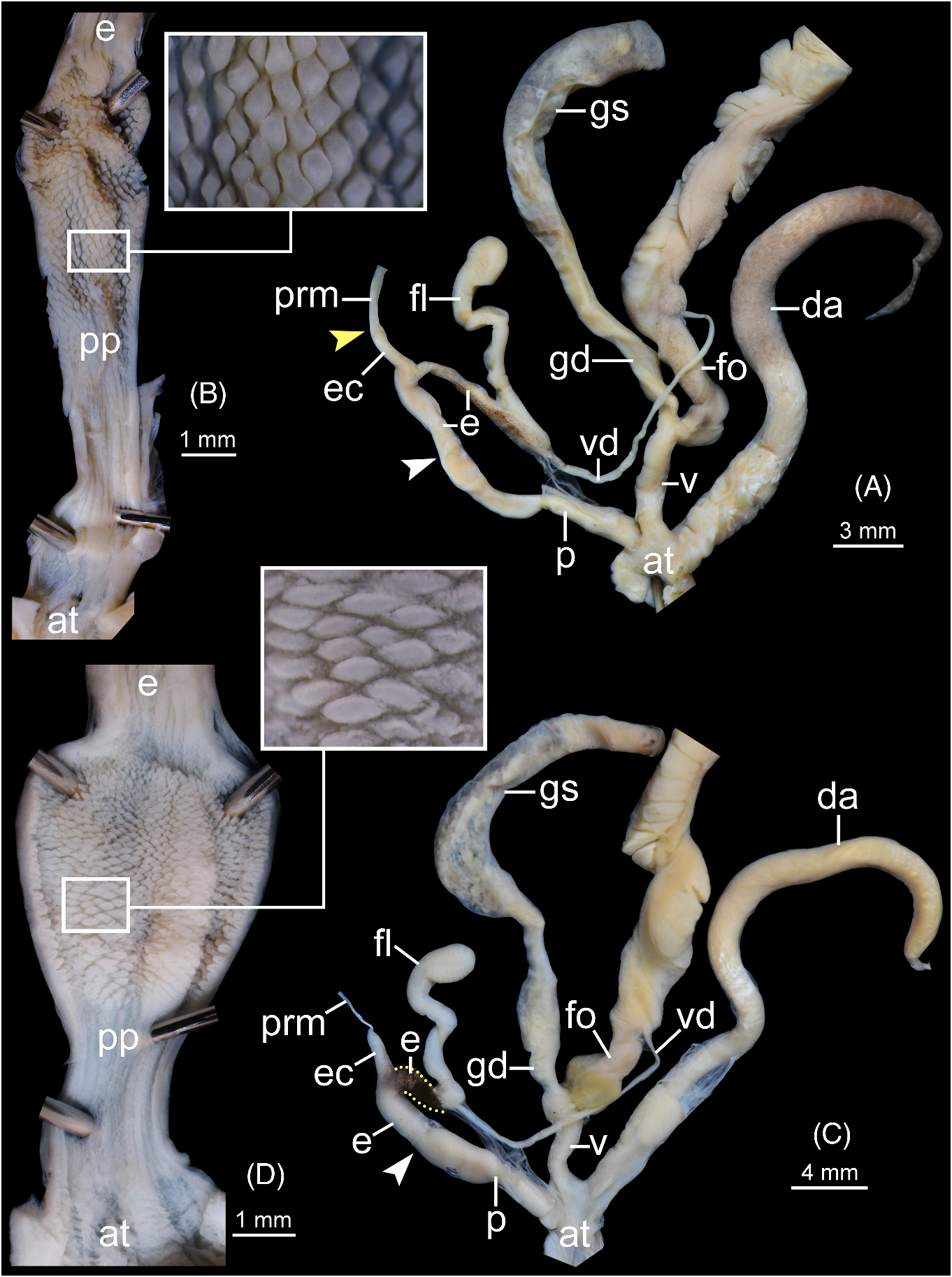
Genital organs ( fig. 4C, D View FIGURE 4 ). Atrium (at) enlarged, very short. Penis (p) elongate, cylindrical, with thin penial sheath covering near atrium. Inner surface sculptured as very finely folded to nearly smooth from atrium to middle of penis, and then gradually transforming to cuboidal penial pilasters (pp). Epiphallus (e) cylindrical, slightly shorter than penis, and distal epiphallus (between epiphallic caecum and flagellum) usually stained with dark colour in adult specimens (see yellow dot lines in fig. 4C View FIGURE 4 ). Epiphallic caecum (ec) straight, smaller diameter than proximal epiphallus, and located at middle of epiphallus. Penial retractor muscle (prm) thin and attached at head of epiphallic caecum. Flagellum (fl) slender, longer than epiphallus, and enlarged at the tip. Vas deferens (vd) thin tube.
Vagina (v) short cylindrical, and about half of penis length. Dart apparatus (da) enlarged and joined to atrium at vagina and penis junction. Gametolytic sac (gs) elongate bulbous (containing one spermatophore inside); gametolytic duct (gd) enlarged cylindrical. Free oviduct (fo) enlarged cylindrical, slightly longer than vagina, and proximally encircled with yellowish or brownish tissue. Oviduct large lobules; prostate gland running alongside oviduct.
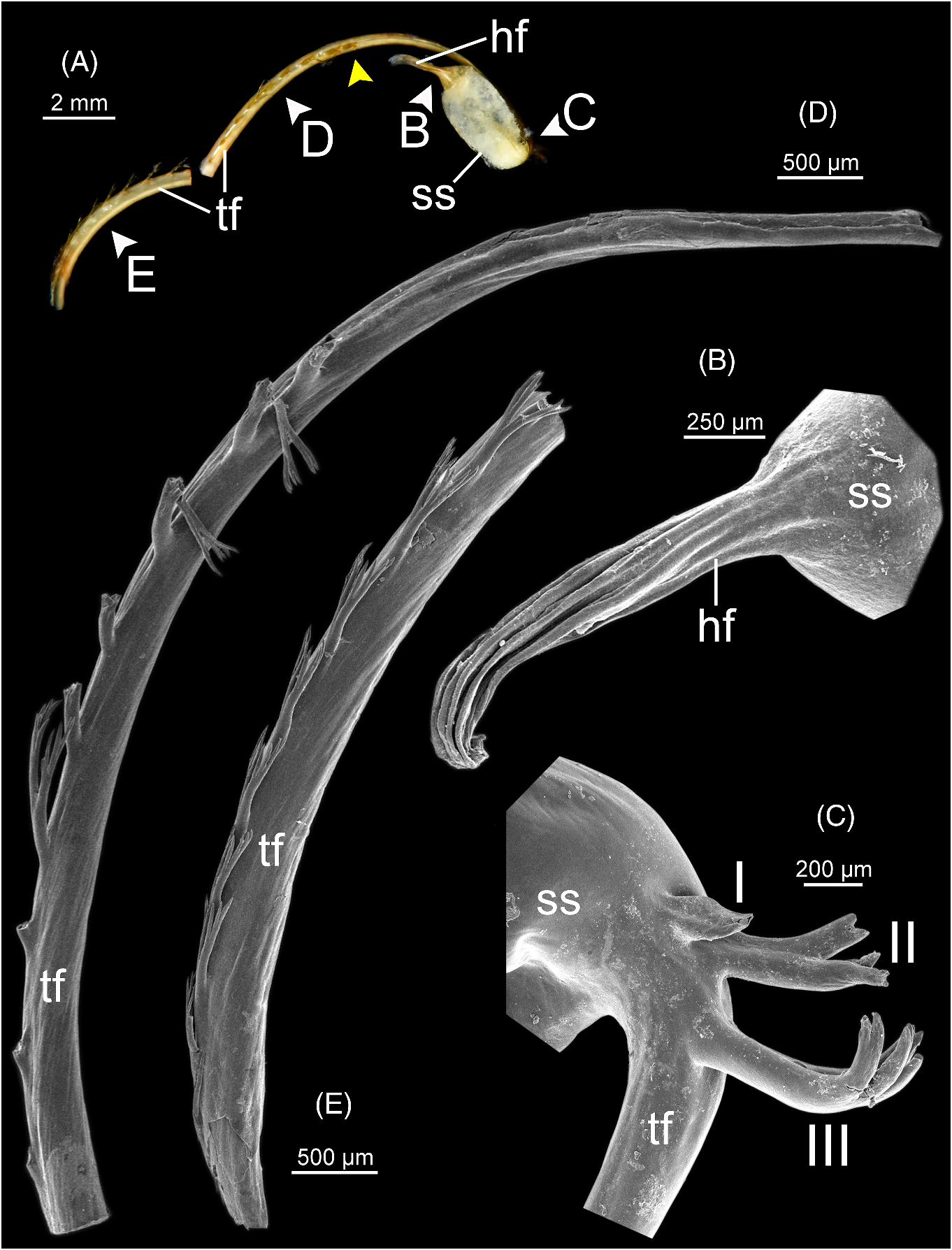
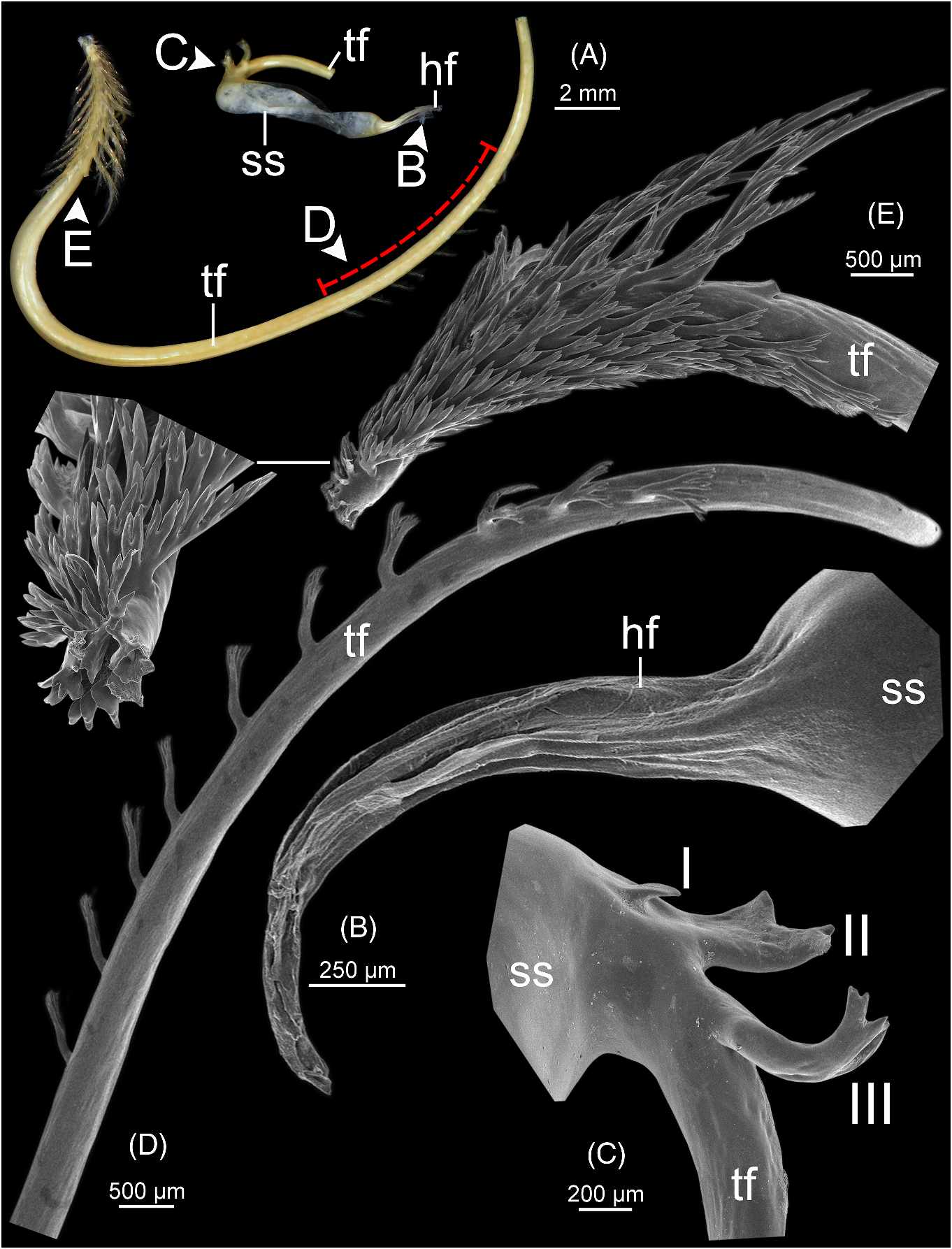
Spermatophore ( fig. 6 View FIGURE 6 ). Spermatophore long and needle-shaped ( fig. 6A View FIGURE 6 ). Sperm sac (ss) enlarged and elongate oval. Head filament (hf) elongate and with smooth longitudinal folds ( fig. 6B View FIGURE 6 ). Tail filament (tf), region close to sperm sac bearing three spines: spine I simple and slender; spines II and III almost similar in size, longer than spine I and with branching into small spinules ( fig. 6C View FIGURE 6 ). Region furthest away without spine; terminal part ca. seven-tenths of its length with series of long branching spines arranged in single row ( fig. 6D View FIGURE 6 ), and then transformed to series of short and delicate branching spines covering terminal part of tail filament ( fig. 6E View FIGURE 6 ).
Radula ( fig. 11B View FIGURE 11 ). Teeth arranged straight with half row formula: 1–(20–21)–72. Teeth shape is similar to those of S. theodori . Central tooth symmetrical tricuspid; lateral teeth asymmetrical tricuspid. Marginal teeth started around tooth number 20 or 21 and with elongate bicuspid.
External features ( figs 2 View FIGURE 2 , 7 View FIGURE 7 ). Living snails with reticulated skin. Eye stalks and body with whitish, creamy-grey to dark grey colour. Four well-developed mantle lobes with same colour as body. Right shell lobe short and left shell absent. Dorsal lobes large and broad: anterior and posterior left dorsal lobes smaller than right dorsal lobe. Caudal fossa and caudal horn present.
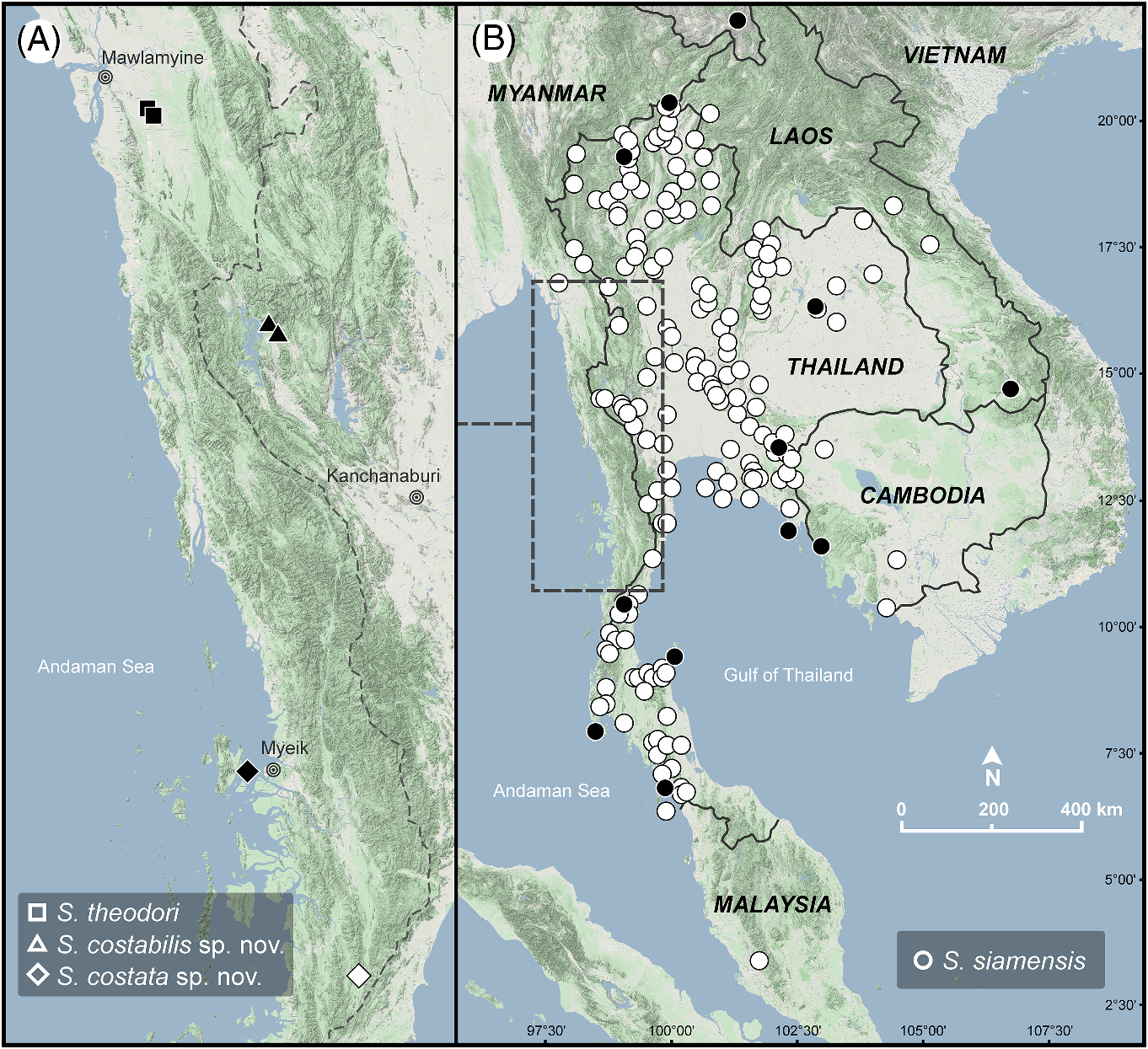
Distribution ( fig. 1 View FIGURE 1 ). Sarika siamensis has a wide distribution, currently known from Thailand, Myanmar, Laos, Malaysia, Singapore, Cambodia, and southern China ( Prasankok & Panha, 2011; Abu-Bakar et al., 2014; Tan et al., 2016; Inkhavilay et al., 2019; Boonmachai & Nantarat, 2020; Dumidae et al., 2020; Sutcharit et al., 2020b). Its distribution in Vietnam and other countries needs further studies. All records of S. siamensis come from sandstone, rocky, limestone, mountainous habitats, and human disturbed areas ( Prasankok & Panha, 2011; Tan et al., 2016; Inkhavilay et al., 2019; Boonmachai & Nantarat, 2020; Dumidae et al., 2020; Sutcharit et al., 2020b).
Remarks. Pfeiffer (1856a) described this species from “ Siam ” [ Thailand] based on specimens in the Cuming collection; however, the type specimen could not be located in the NHM, London collection. This species has been classified to the Indian genus Cryptozona since the classification of Panha (1996); however, its generic position has been uncertain. Most studies on this species have concentrated at the population level because this species is the most widespread species in mainland Southeast Asia ( Prasankok & Panha, 2011; Tan et al., 2016; Inkhavilay et al., 2019; Boonmachai & Nantarat, 2020; Dumidae et al., 2020; Sutcharit et al., 2020b) and is currently reported as the intermediate host for the rat lungworm that causes eosinophilic meningitis in human ( Vitta et al., 2016; Dumidae et al., 2020).
There were no records of S. siamensis in Myanmar until our survey in 2016, where a S. siamensis population was discovered in an urban area (Hpa-An City, Myanmar), but we could not find them in suburban or rural areas. In addition, Macrochlamys kelantanensis Möllendorff, 1902 , a widespread species in Malaysia and Thailand, was also found at same locality with S. siamensis (Pholyotha et al., 2020b) . Thus, we consider that this population of S. siamensis has recently expanded its range into Myanmar, possibly associated with anthropogenic activities due to the high mountain barrier to snail dispersal covering from northern Thailand and extending along the Myanmar border ( Gupta 2005). An accidental introduction through horticultural trade activities or association with human activities has been reported in Singapore ( Tan et al., 2016), Chintamanis in Malaysia ( Prasankok & Panha, 2011), Yunnan in China ( Sutcharit et al., 2020b), and Kampong Speu in southern Cambodia ( Sutcharit et al., 2020b).
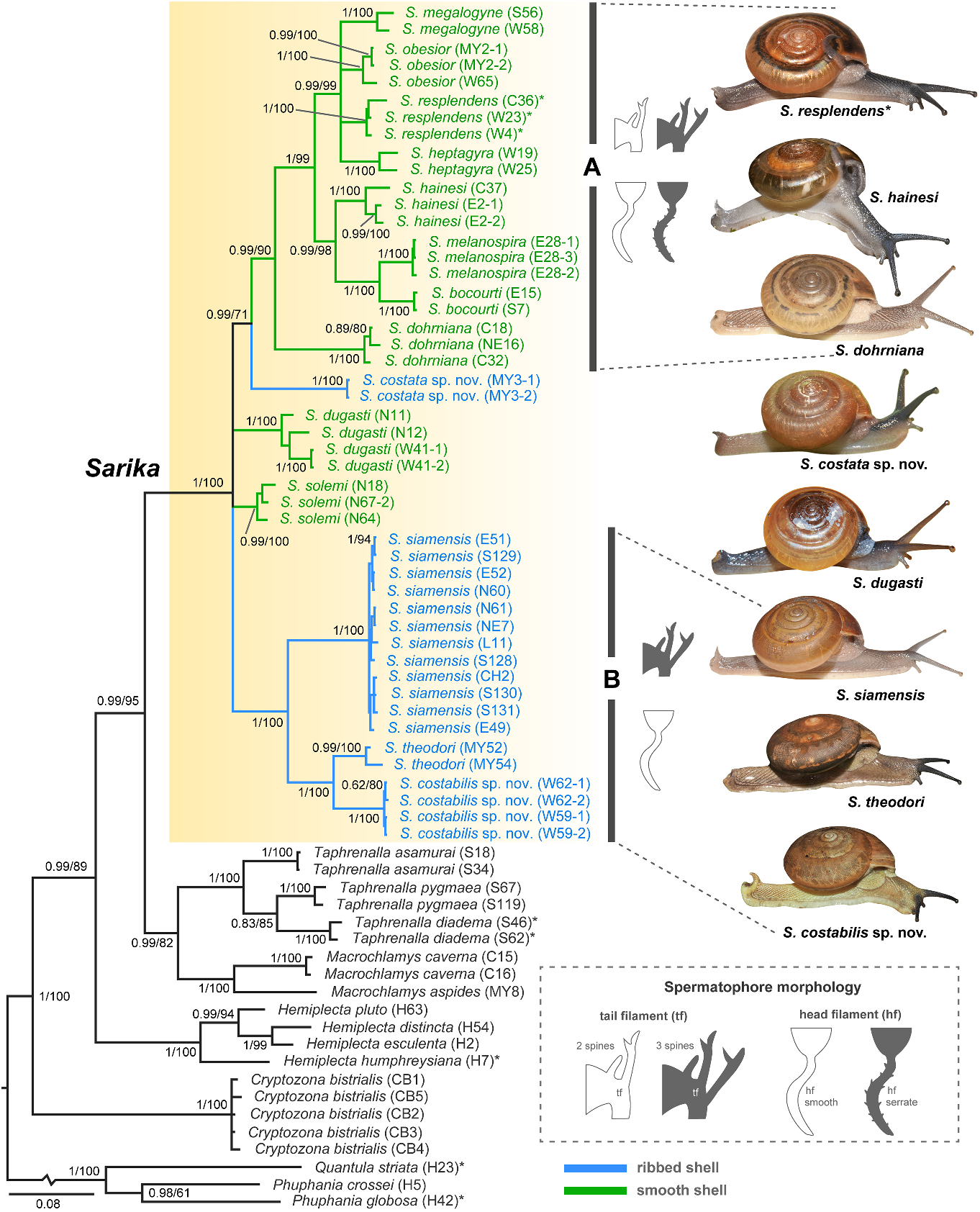
In addition, S. siamensis exhibits a rather wide range of shell colour variation from monotone whitish ( fig. 7B, C View FIGURE 7 ) or two-tone, much paler beneath ( fig. 7A, D View FIGURE 7 ), and with or without a peripheral band ( fig. 7C, D View FIGURE 7 ). This species also exhibits a rather wide range of body colour variation from whitish ( fig. 7A View FIGURE 7 ) to creamy-grey ( fig. 7C View FIGURE 7 ) or grey ( fig. 7B, D View FIGURE 7 ). In the past, there were some examples of taxonomic problems from these variations of S. siamensis . A white unbanded variety with faintly decussated shells was erroneously recorded as S. hainesi (Marten, 1860; Marten, 1867; Tryon, 1886), while a large, pale yellowish shell with a reddish band was also erroneously described as S. birmana (Pfeiffer, 1857) (Marten, 1860; Marten,1867; Morelet, 1875). In this study, we examined all the variations in S. siamensis and found that the genitalia of these morphs were identical. In addition, our molecular phylogeny also supported that all these morphs were grouped together within the S. siamensis clade ( fig. 2 View FIGURE 2 ).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Sarika siamensis (Pfeiffer, 1856)
| Pholyotha, Arthit, Sutcharit, Chirasak, Lin, Aung & Panha, Somsak 2022 |
Naninia [sic] mouhoti:
| Hemmen, J. & Hemmen, C. 2001: 45 |
Cryptozona siamensis:
1996Hemiplecta siamensis:
| Hemmen, J. & Hemmen, C. 2001: 44 |
| Panha, S. 1996: 34 |
| Solem, A. 1966: 27 |
Xestina siamensis:
| Tomlin, J. R. le 1932: 319 |
| Kobelt, W. 1905: 1136 |
Nanina (Xesta) siamensis:
| Fischer, H. & Dautzenberg, P. 1904: 394 |
Nanina mouhoti:
| Morlet, L. 1889: 124 |
| Tryon 1886: 85 |
Nanina siamensis:
| Tryon 1886: 86 |
| Martens, E. von 1867: 71 |
Helix (Nanina) mouhoti
| Pfeiffer, L. 1868: 116 |
| Pfeiffer, L. 1860: 136 |
Helix siamensis Pfeiffer, 1856a: 32
| Pfeiffer, L. 1856: 32 |
Helix siamensis: Pfeiffer, 1856b: 76
| Morelet, A. 1875: 247 |
| Pfeiffer, L. 1859: 60 |
| Pfeiffer, L. 1856: 76 |