
Rhynchelmis paraolchonensis, Giani & Martinez-Ansemil, 1984, Giani & Martinez-Ansemil, 1984
|
publication ID |
https://doi.org/10.5281/zenodo.194252 |
|
DOI |
https://doi.org/10.5281/zenodo.6201863 |
|
persistent identifier |
https://treatment.plazi.org/id/03C5878E-FFF0-D41D-B2CF-FF197EC96ECC |
|
treatment provided by |
Plazi |
|
scientific name |
Rhynchelmis paraolchonensis |
| status |
|
Pseudorhynchelmis paraolchonensis View in CoL ( Giani & Martinez-Ansemil, 1984)
( Fig. 10 View FIGURE 10. P )
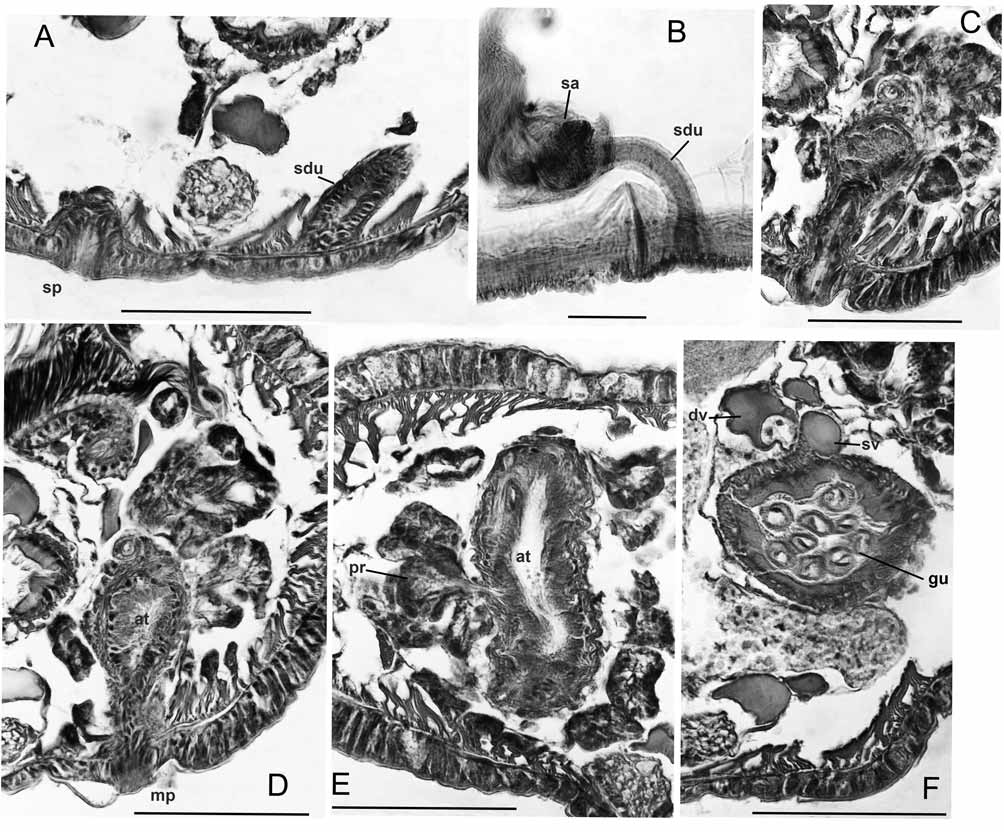
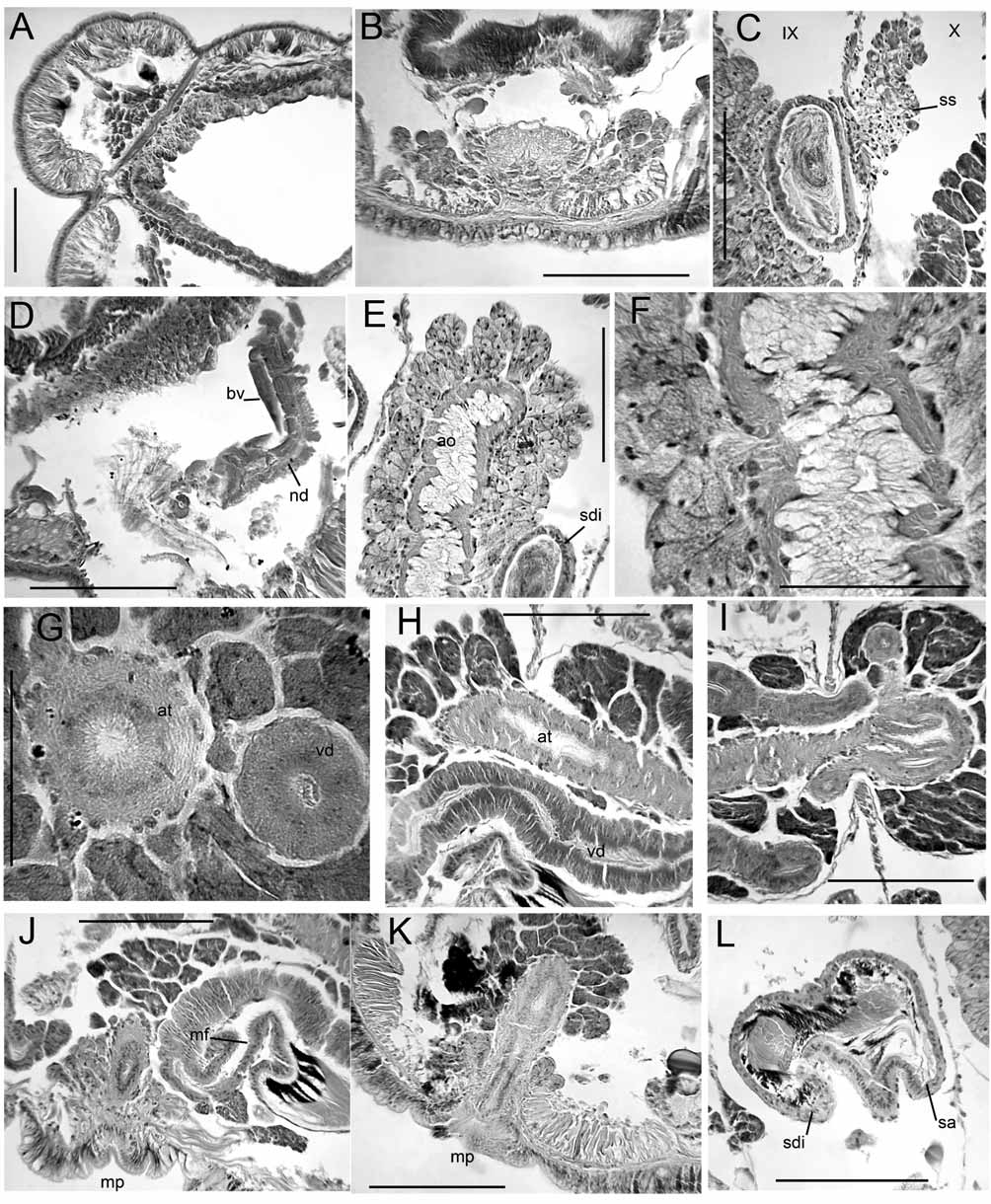
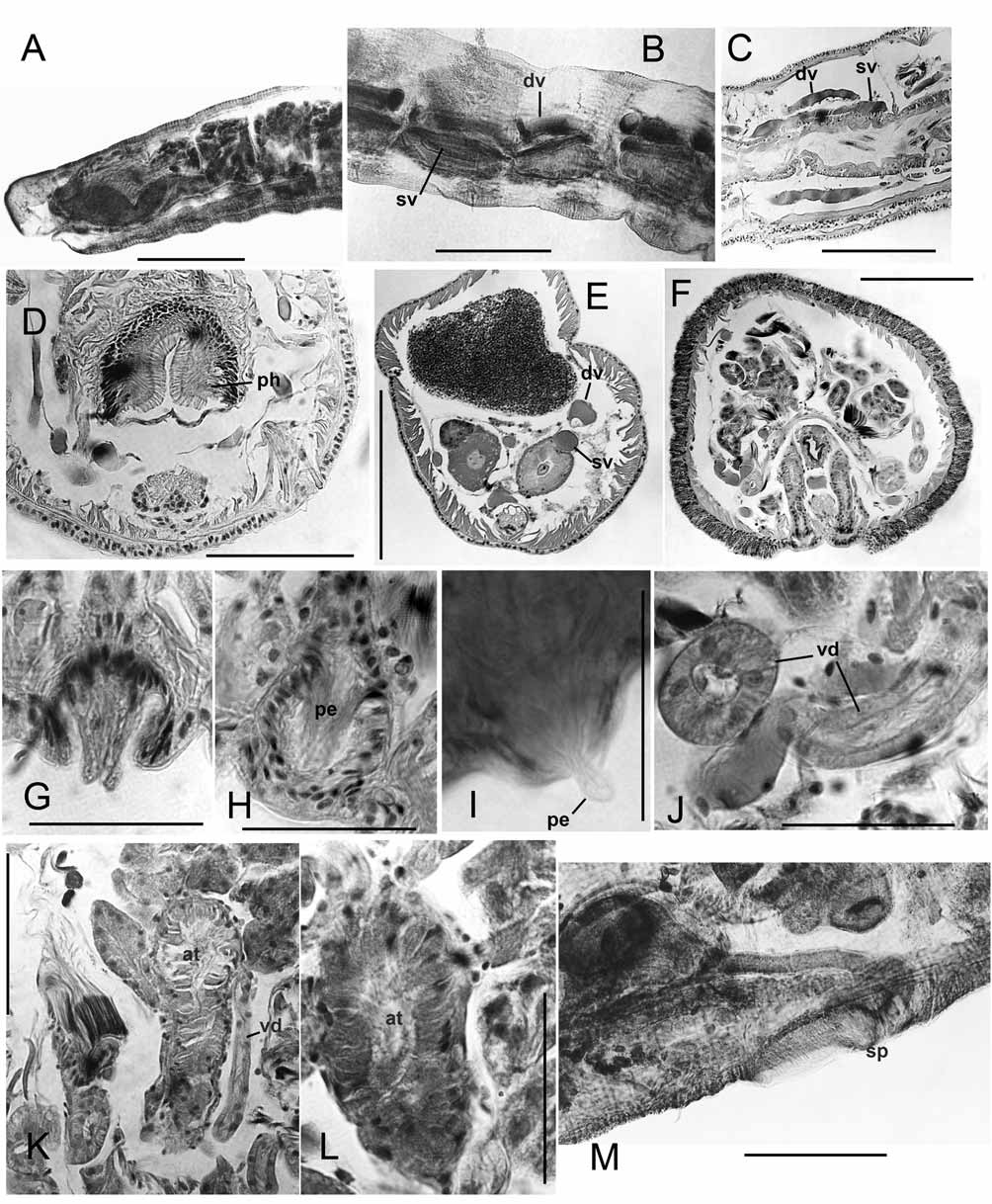
Supplementary description. Dorsal pad of pharynx only moderately developed. Longitudinal muscles in weak bands that are not curled at margins; oblique dorsolateral muscles not strongly developed. A single pair of convoluted commissural vessels in each anterior segment may enter adjacent segment; commissures absent in segments posterior to XI, but a loop enters the sperm sacs from X. Dorsal and ventral vessels prominent, with connections to perivisceral sinus; perivisceral sinus forms a supra-intestinal vessel in posterior segments, without cardiac cells ( Fig. 10 View FIGURE 10. P F). Nephridia may begin in XIII, but absent in many segments. Nephridia with small postseptal expansion; convoluted tubules extend dorsally, but may also follow ventral blood vessel through several segments; duct only slightly widened at inconspicuous nephropore.
Spermathecae with irregular, sacciform ampulla that may enter XI, but does not connect to gut; ampulla contains loose, unordered sperm. Spermathecal duct a long, narrow cylinder, length more than 5 times width; duct slightly widened before pore, but otherwise not modified ( Figs. 10 View FIGURE 10. P A–B).
Atrial muscle layer to nearly 10 Μm thick, lining cells moderately thick, columnar. Prostates in large, irregular bundles; bundles are loosely packed ( Figs. 10 View FIGURE 10. P D–E). Both pairs of vasa deferentia functional; diameter about 20 Μm (20–27) near funnels, narrowing very slightly to about 18 Μm (15–22); lining cells cuboidal, not glandular. Both vasa join atrium near ental end, travelling at most a short distance under muscles. Male pore on a small papilla within a shallow depression; epidermis slightly thickened, but muscles not strongly developed ( Figs. 10 View FIGURE 10. P C–D).
Remarks: Pseudorhynchelmis
Kaygorodova & Liventseva (2007) expanded this genus to include ten taxa, including an undescribed species, but some of these species are in need of revision, and at least one is probably invalid ( Martin & Kaygorodova 2008). There is little morphological evidence to link these species with Rhynchelmis , other than the relative positions of the spermathecae and atria. The same configuration occurs in other accepted genera ( Tatriella Hrabĕ and Secubelmis Fend & Gustafson ), and in at least three undescribed Nearctic species that may represent additional lineages (S. Fend, unpublished). Other configurations occur in multiple genera; for example, the spermathecae are in the first preatrial segment in the semiprosoporous genera Stylodrilus Claparède , Eclipidrilus Eisen and Tenagodrilus Eckroth & Brinkhurst. Inclusion in Rhynchelmis , based on the position of the first spermathecae in VIII, would almost certainly result in a polyphyletic genus, and would also require the equally unsupported inclusion of Tatriella and Secubelmis . The recent decisions by Kaygorodova & Liventseva (2007) and Martin & Kaygorodova (2008) to separate these taxa from Rhynchelmis on both morphological and molecular grounds are reasonable from a phylogenetic view, and retention of Pseudorhynchelmis is the best solution for conserving the present taxonomic system.
While the species assigned to Pseudorhynchelmis appear to form a (phenetic) group based on overall similarity, there are few, if any, conclusive synapomorphies distinguishing them from other lumbriculid genera. This problem is evident in the recent numerical analyses by Kaygorodova & Liventseva (2007): the species do form a cluster in the neighbor-joining tree, but were not resolved as a monophyly in a maximum parsimony tree. Martin & Kaygorodova (2008) listed three possible synapomorphies, two of which are not shared by all members of the group: (1) vasa deferentia much wider entally than ectally (noted by Hrabĕ 1982), (2) atrial lining cells thick and glandular, and (3) pendant penes in penis sacs. Based on our limited sampling of the group, additional potential apomorphies for the Baikal group include the distinct pharyngeal pad, the supra-intestinal blood vessel, and the penes formed by extrusion of atrial lining cells, but these characters are not unusual within the family, and have to be verified on more species. Even allowing for reversals within some characters, defining the genus on the basis of these apparent synapomorphies may require the exclusion of the western European P. paraolchonensis (see below).
A loop of the posterior vas deferens into the post-atrial segment is well documented (e.g., Giani & Martinez-Ansemil 1984, Martin et al. 1998, Semernoy 2004), and distinguishes Pseudorhynchelmis species from all Rhynchelmis , but as this is assumed to be a plesiomorphic character state ( Hrabĕ 1984), it cannot be used to define Pseudorhynchelmis ( Martin & Kaygorodova 2008). Pseudorhynchelmis atria are generally shorter than those of Rhynchelmis , usually extending at most into the ovarian segment, and in some cases they are more an elongate pear-shape than tubular. In the few Rhynchelmis species having atria commonly limited to X and XI, the atria are convoluted, and would extend through more posterior segments if straight ( Martin et al. 1998, Fend & Brinkhurst 2000). Nevertheless, Pseudorhynchelmis atria appear to represent a more general state within the family than the long atria of Rhynchelmis .
The suggestion by Martin & Kaygorodova (2008) that “pendant penes” may be a synapomorphy for Pseudorhynchelmis may be somewhat refined by the present observations. The presence of elongate cells lining the atrial duct at the male pores of P. alyonae and P. shamanensis (protruding in the latter) suggest penes formed by extrusion of atrial lining cells (type 2 penes, sensu Cook 1967). Figure 2 View FIGURE 2. R B in Martin & Kaygorodova (2008) suggests that similar penes also occur in Pseudorhynchelmis parva (Michaelsen), and illustrations by Semernoy (2004, Figs. 225, 227, and 234) may show the same penial structure in other Pseudorhynchelmis species. This character may be difficult to observe in most preparations, and needs confirmation in well-fixed specimens of other species. Although various types of penes occur in Rhynchelmis (e.g., Fend & Brinkhurst 2000 for the Nearctic species), their basic structure differs from that of these Pseudorhynchelmis species.
The atrial histology of at least some Pseudorhynchelmis differs from that of Rhynchelmis . The lining cells of the observed species, plus those observed by Martin & Kaygorodova (2008), are thick and glandular, but based on their descriptions, P. spermatochaeta and P. minimaris may be exceptions ( Semernoy 1982, 2004). The prostate bundles in the material observed here are relatively larger, and more irregular in size, shape, and placement than those of Rhynchelmis , in accordance with the observations by Martin & Kaygorodova (2008) and Kaygorodova & Liventseva (2007).
Spermathecae of Pseudorhynchelmis species do not resemble those of any of the Rhynchelmis groups (see also Giani & Martinez-Ansemil 1984, Semernoy 2004, Martin & Kaygorodova 2008). Unlike R. ( Rhynchelmis ), the Sutroa group, some of the Rhynchelmoides group, and R. brooksi / malevici , the ampullae are rather simple sacs, without diverticula or obvious ducts to the gut. In contrast to those Rhynchelmoides group species lacking a spermatheca-gut connection, the ampullae are not divided into histologically distinctive sections, with viable sperm concentrated in the ectal region. The ectal ducts do not resemble those of any of the Nearctic Rhynchelmis ; and are typically more narrowly tubular than in R. ( Rhynchelmis ) (e.g., Fig. 9 View FIGURE 9. P M vs. 3I). The spermathecal ampulla joins the gut via a short, but distinct ental duct in those Rhynchelmis species having a spermatheca-gut connection (e.g., Fig. 3 View FIGURE 3. R K). In contrast, Semernoy’s (2004) illustrations of several Baikal Pseudorhynchelmis species (not available for study here) show an inconspicuous median junction, which may need to be verified with sectioned material. Martin & Kaygorodova (2008) questioned the validity of the spermatheca-gut connection in these taxa; although Kaygorodova & Liventseva (2007) assumed that it was correct for purposes of analysis, they defined the genus in part by its absence.
The structure of the pharynx is usually not described in detail in lumbriculids. The two observed Baikal Pseudorhynchelmis (but not paraolchonensis ) had a strongly developed dorsal pad in II–III or IV, whereas this did not occur in any observed R. ( Rhynchelmis ), or in R. orientalis . Semernoy (1984, Figs. 233 and 234) illustrates pharyngeal pads in two other Pseudorhynchelmis species. The dorsal wall of the pharynx was only slightly developed, and both dorsal and ventral walls were eversible in other Rhynchelmoides and Sutroa group taxa ( Fend & Brinkhurst 2000).
No mention of the blood vessels was made in the original descriptions of P. alyonae and P. shamanensis , and the vascular system was not completely visible in the available material. However, our observations suggest that these species differ from most Rhynchelmis species in the complete lack of posterior lateral blood vessels, the anterior commissures with a strong anterior loop, and the supra-intestinal vessel formed from the perivisceral sinus. Nephridia were difficult to see clearly, as no sagittally sectioned specimens were available. Although taxonomically important, nephridia and blood vessels are difficult to see in most preparations, and are not described for many lumbriculids.
The lack of a proboscis, the lack of a spermathecal-gut connection, and the testes with functional vasa deferentia in XI support the removal of P. paraolchonensis from Rhynchelmis , but as these are likely plesiomorphic states, they only indirectly support its placement in Pseudorhynchelmis. The blood vessels, the somewhat median placement of genital pores, the posterior vas deferens entering XI, the structure of the spermathecae, and the histology of the atrium also suggest affinity with P. olchonensis and similar species. Giani & Martinez-Ansemil (1984) differentiated P. paraolchonensis from P. olchonensis by the position of the clitellum, number of segments, and the posterior vas deferens length not greater than that of the anterior. Martin et al. (1998) noted the distinctive chaetae, which were illustrated in the original description as being proportionally much thicker than in other members of the genus, and also noted the short atria and greater overall size. Pseudorhynchelmis paraolchonensis differs from at least some of the Baikal species in lacking potential synapomorphies for Pseudorhynchelmis: the ectal end of the atrium forms a slight papilla, but the lining cells do not appear modified; the vasa deferentia are not strongly tapered; and the pharynx does not have a strongly developed dorsal pad.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Rhynchelmis |