
Ramiellona strigosa setosa Righi, 1972
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3753.6.3 |
|
publication LSID |
lsid:zoobank.org:pub:8DF3EFAA-F8CC-4D4C-A830-D07D76C2D1C0 |
|
DOI |
https://doi.org/10.5281/zenodo.5616743 |
|
persistent identifier |
https://treatment.plazi.org/id/FD1F8789-9A48-FF89-1DEC-A823FDC8FB05 |
|
treatment provided by |
Plazi |
|
scientific name |
Ramiellona strigosa setosa Righi, 1972 |
| status |
|
Ramiellona strigosa setosa Righi, 1972
Figures 7 View FIGURE 7 , 8 View FIGURE 8 , 9 View FIGURE 9
Ramiellona strigosa setosa Righi 1972: 222 , Figs 23–28.
Localities. Mexico. (1) Chiapas state, Chajul, Municipality of Marquez de Comillas, 1 km S from town, in fallows and tropical rain forests over ferrallitic soils at 0–50 cm depth. 16°06’24”N, 90°55’24”W, 190 m a.s.l.: 11/14/1981, 11/15/1981, C. Fragoso and P. Lavelle; 05/1,3,5/1982, C. Fragoso; 06/8,9/1982, P. Lavelle; 07/13,14/1982, 09/9,10/ 1982, 11 /09/1982, 12/30,31/1982, 5/5,6,16/1983, C. Fragoso. - 2 km SE from town, disturbed tropical rain forest with seedlings of cocoa over ferrallitic soils at 0–50 cm depth. 16°06’42”N, 90°54’48”W, 200 m a.s.l., 10/29/1983, 12/29/1983, 02/01/1984, 05/03/1984, 08/28/1984, 12/15/1984, C. Fragoso. - 2 km SW from town close to the river Lacantun, 10 years pasture, within soil at 0–50 cm depth. 16°06’30”N, 90°55’30”W, 200 m a.s.l., 10/31/1983, 11/ 01/1983, 02/16/1984, 05/05/1984, 08/27/1984, 12/16/1984, C. Fragoso. - 3 km SW from town, very near to the river Lacantun in a recently deforested parcel over alluvial soils, within soil. 16°06’15”N, 90°57’00”W, 200 m a.s.l., 12/12,14/1984, C. Fragoso. - In front of town and crossing the river Lacantun, 2 km NW inside Montes Azules Biosphere Reserve, tropical rain forest over alluvial soils at 0–50 cm depth. 16°07’24”N, 90°56’24”W, 180 m a.s.l., 11/16/1981, C. Fragoso and P. Lavelle; 06/5,6,7/1982 P. Lavelle, 04/29/1982, 05/1,4,6,/1982, 07/10,11,12/ 1982, 08/9,10/1982, 09/9,10/1982, 12/28,29/1982, 01 /01/1983, 05/5,7/1983, 11 /02/1983, 05/4,6/1984, 08 /29 / 1984, 12 /17/1984, 01 /01/1986, C. Fragoso. (2) Chiapas state, Miramar, Municipality of Ocosingo. Near to the lagoon inside Montes Azules Biosphere Reserve; tropical rain forest, within soil. 16°22’42”N, 91°16’43”W, 300 m a.s.l., 08/14,15/1976, 10/14,15/1976 P. Lavelle. (3) Chiapas state, Bonampak, Municipality of Ocosingo. SW from the archaeological site; tropical rain forests over pH neutral rendzina soils at 0–50 cm depth. 16°42’07”N, 91°03’52”W, 300 m a.s.l., 08/25/1977, 11/19/1977, P. Lavelle; 11/11/1981, C. Fragoso and P. Lavelle. N from the archaeological site; tropical rain forests over clay soils at 0–20 cm depth. 16°42’48”N, 91°03’50”W, 340 m a.s.l., 11/12/1981, C. Fragoso and P. Lavelle. - Archaeological site; grass over clay soils at 0–20 cm depth. 16°42’13.7”N, 91°03’53.7”W, 300 m a.s.l.. 11/11/1981, C. Fragoso and P. Lavelle. (4) Chiapas state, San Cristobal de las Casas. SW outside town, Fraccionamiento Articulo 115, in front of ECOSUR Research buildings; hill covered with slightly disturbed pine-oak forest, within soil at 20 cm depth. 16°42’1.9”N, 92°36’47.1”W, 2130 m a.s.l.. 11/29/ 1997, C. Fragoso and J. Bueno. (5) Chiapas state, Ixhuatan. Road 195, 4 km after town, edge of road and outside tropical forest 17°19’21.3”N, 93°0’2.3”W, 350 m a.s.l., 11/29/1997, C. Fragoso and J. Bueno. (6) Tabasco state, Municipality of Cardenas, Town C27 Ing. Eduardo Chavez Ramirez, taking the road 180, 28 km before town of Cardenas coming from Coatzacoalcos; upper 20 cm of soil of a cocoa plantation. 18°01’29.7”N, 93°39’21.7”W, 20 m a.s.l., 07/14/1984, C. Fragoso and P. Lavelle. (7) Tabasco state, Municipality of Huimanguillo. Close to a forest plantation of Tectona grandis View in CoL , in pastures at 0–10 soil depth, 40 m a.s.l.. 03/11/1997, C. Castro. (8) Tabasco state, Agua Blanca, Ejido Las Palomas, Municipality of Macuspana. Within the tropical forest that surrounds the waterfalls, under decaying logs and within soils. 17°37’18.1”N, 92°28’25.1”W, 120 m a.s.l.. 12/20/1991, C. Fragoso and P. Rojas; 12/12/2006, C. Fragoso and P. Rojas.
Material examined. Chajul: 24 clitellates, 5 dissected, IEOL 1932, 1945, 1978–3, 1962–1, 2324, 19 externally revised, IEOL 1962, 1940, 1978; Miramar: two clitellates, one dissected, IEOL 1915; Bonampak: 7 clitellates, two dissected, IEOL 2450, 2461, 5 externally revised, IEOL 1981, 2450; San Cristobal de las Casas: two clitellates, one dissected, IEOL 4040; Ixhuatan: one clitellate dissected, IEOL 4090; Huimanguillo: one clitellate dissected, IEOL 3313–1; Agua Blanca: one clitellate externally revised, IEOL 4239, one aclitellate adult dissected, IEOL 1979.
Description. In the following we present characters that were found invariable in the examined populations; in the section entitled "Variations" we present some external characters that varied within and between populations.
External. Pigment absent. Prostomium prolobous. Peristomium with longitudinal grooves, also present in second and third segment. Secondary annulations in preclitellar and postclitellar segments. Setae eight per segment, closely paired in anterior region, widely paired in posterior region. Conspicuous and robust paired dimorphic penial setae (a always larger than b) in 17 and 19 ( Figs 8 View FIGURE 8 , 9 View FIGURE 9 ); seta a with abundant ornamentation, more evident (like teeth) in the distal extreme which ends as a thin filament ( Figs 8 View FIGURE 8 A,B, 9A,B); seta b with a spoon-like apex, and with less and smaller ornamentations ( Figs 8 View FIGURE 8 C,D, 9C,D). Setae a and b of 18 not visible in the majority of the examined individuals. Genital setae absent. Clitellum saddle-shaped. Large dorsal pores present all along the body, first functional pore in 12/13. Female pores in 14, presetal slightly median to A. Two pairs of prostatic pores in 17 and 19 just at base of penial seta. Spermathecal pores paired in 7/8 and 8/9, centered in AB. Genital marks mid-ventral and unpaired as intersegmental ovoidal-rectangular papillae; small and paired diagonally oriented marks, generally in AB and always intraclitellar.
Internal. Septa 5/6, 12/13 and 13/14 thin and membranous, 6/7–11/12 muscular, funnel-shaped, imbricated and joined by several dorsal and lateral connective-muscular tissue fibers. One large gizzard in 5, extending 2–3 segments backwards. Extramural calciferous glands absent. Esophagus with internal lamellae covering the entire esophageal wall in segments 8–12; larger in segments 9–11. Origin of intestine in 14/15. Dorsal typhlosole starting in 16 or 17, laminar and small, increasing abruptly in size in 22 or 23 with free edge divided into three deep ridges (trifid) until its end, tens of segments before anus. Typhlosole with lateral folds, dorso-ventrally oriented in segments 23–28, 29. Smaller dorso-lateral typhlosoles in the region of segments 21–29, at both sides of main typhlosole. Intestinal caeca present as 3–4 pairs of dorsal pouches on the intestine of segments 23–27; some segments with apparently more than two caeca, due to the invasion of posterior caeca.
Single dorsal vessel visible throughout; two pairs of lateral commissures per segment dorsally joined to the intestine in 15–17. Ventral vessel present. Large latero-ventral vessels running forward under the gizzard. Supraesophageal vessel visible in 7–12. Lateral hearts in 7–10; latero-esophageal hearts in 11 and 12. Paired infraesophageal vessels in 7–12, which join supra-esophageal vessel through lateral esophageal commissures. Extra parietal ventral vessels in each side of segments 13, 14 and 15, running outside of male gonoduct. Paired clusters of tufted microphridia in ventral parietes of segments 2–5. Septal nephridia in preclitellar segments. Parietal, closed meronephridia extending backwards from 14, 8–16 per segment (4–8 on each side). The median-ventral nephridium of last segments larger and with small nephrostome in the preceding segment.
Holandric. Testes and iridescent male funnels of 11 larger than those of 10. Male gonoducts muscular from segment 14, entering body wall in equator of 17. Two pairs of acinous seminal vesicles in 9 and 12, the anterior pair smaller than the posterior one. Two pairs of tubular prostates in 17 and 19, strongly coiled and fixed to the intestine and septa by connective tissue; the glandular part wider and 4–5 times longer than the muscular duct. Follicles of penial setae a and b joined by its ectal end, forming a muscle pouch; these pouches fixed by muscular stripes to the lateral wall of the body. Each follicle with two undeveloped setae located in a separated pouch. Ovaries in 13. Two pairs of similar-sized spermathecae discharging in 7/8 and 8/9, ampulla and duct in posterior segments (8 and 9, respectively), duct wider and shorter than elongate ampulla, the latter with sausage shape. Diverticle in anterior segments (7 and 8), sessile, flat, ovoid and fixed to the walls, with several small seminal chambers.
Variations. Length: Chajul: 140–175 mm (contracted), 190–300 mm (relaxed); Miramar: 185 mm (relaxed); San Cristobal: 80–100 mm (contracted); Ixhuatan: 95 mm; Huimanguillo: 200 mm; Agua Blanca: 120 mm (contracted).
Width: Chajul: 3.6–6.1 mm; Miramar: 5 mm; San Cristobal: 5.3 mm; Ixhuatan: 4.4 mm; Huimanguillo: 4.3 mm; Agua Blanca: 5.4 mm.
Number of segments: Chajul: 243, 237; San Cristobal: 210; Ixhuatan: 206; Huimanguillo: 328.
Clitellum: Chajul: 14–18 (10 ind.) ( Fig. 7 View FIGURE 7 A), 1/2 13 –18 (one ind.), 13–19 (one ind.); Bonampak: 14–18 (4 ind.), 13–20; Miramar: 1/2 13 –18, 14–20; San Cristobal: 14–18 ( Fig. 7 View FIGURE 7 E); Ixhuatan: 14–18; Huimanguillo: 1/4 13 – 18 ( Fig. 7 View FIGURE 7 D); Agua Blanca: 14–18 ( Fig. 7 View FIGURE 7 C), 1/2 13 –19, 1/2 20.
Genital marks: Chajul ( Fig. 7 View FIGURE 7 A): Unpaired midventral (in the following: UM) present in some of the intersegments 10/11–14/15, 19/20–21/22; paired (in the following: P) in some of the intersegments 14/15–18/19; the marks of 14/15 can be paired or unpaired, this varying in each individual. In some individuals transverse rows of circular areas within the marks of 10/11, 12/13–15/16 ( Fig. 7 View FIGURE 7 B). Bonampak: UM in some of the intersegments 9/ 10, 10/11, 12/13, 14/15, 19/20, 20/21; P in some of the intersegments 13/14–16/17; the marks of 14/15 can be paired or unpaired, this varying in each individual. Miramar: UM in some of the intersegments 12/13–14/15, 19/ 20–21/22; P in 15/16, 16/17. San Cristobal: UM in some of the intersegments 10/11, 12/13–14/15, 20/21–24/25; P in 15/16, 16/17 ( Fig. 7 View FIGURE 7 E). Ixhuatan: UM in 8/9–10/11, 14/15, 15/16, 20/21–23/24; P in 16/17. Huimanguillo: UM in 10/11, 13/14, 14/15, 19/20, 21/22, 22/23; P in 15/16–18/19; transverse rows of circular areas within the mark of 15/16 ( Fig. 7 View FIGURE 7 D). Agua Blanca: UM in 10/11–13/14, 20/21, 21/22 ( Fig. 7 View FIGURE 7 C).
Penial setae ornamentation: Chajul, Miramar, San Cristobal: serrations limited to the last distal third, before apex, small, abundant and surrounding entire circumference of setae; at the apex thorns conspicuous and scarce ( Fig. 8 View FIGURE 8 B). San Cristobal: very similar, but apex thorns much scarcer. Ixhuatan: the single individual with apex of and the original descriptions of Ramiellona strigosa strigosa Gates, 1962 from Guatemala, Ramiellona eiseni
( Michaelsen, 1911) from Guatemala, and Ramiellona strigosa setosa Righi, 1972 from Mexico.
both a and b setae very different from all populations, ending not as a spoon or as a tip, but twisted and truncated ( Fig. 9 View FIGURE 9 F,H). Huimanguillo and Agua Blanca: serrations and thorns abundant and conspicuous in both a and b setae ( Fig. 9 View FIGURE 9 B,D).
Remarks. Ramiellona strigosa strigosa was described by Gates (1962) from six adults, five aclitellate and one clitellate. The material had been collected by Eisen in Guatemala, in the highland pine forest region of Totonicapan, during his travel across the western and southern regions of Guatemala ( Eisen 1903). It remained at the California Academy of Sciences until its revision by Gates, sixty years later. Gates (1962) pointed out the similarity with R. eiseni —collected also by Eisen in the proximities of Huehuetenango during the same trip, and described by Michaelsen (1911) from a single clitellate adult. Several differences indicated by Gates for both species were found in the material of R. strigosa setosa revised in this study, varying between populations and even within populations (e.g. number of genital marks, presence or absence of transverse circular areas within each marking, extension of clitellum). Unfortunately Michaelsen (1911) did not record other internal characters that would support (or not) the separation of both species. By now only the number of seminal vesicles and the presence/absence of calciferous lamellae separate both species ( Table 1 View TABLE 1 ). Later on Righi (1972) described the subspecies R. strigosa setosa from Chiapa de Corzo. Although this author did not indicate on which differences the erection of the subspecies was based, we can conclude from Table 1 View TABLE 1 that the shape of penial setae, position of seminal vesicles and conspicuous posterior somatic setae led Righi to erect a new subspecies; it was considered as R. strigosa by the shape of the spermathecae and the presence of calciferous lamellae in segments 8–12. This subspecies shares a key feature with all the populations studied in this paper: dimorphic penial seta a and b with different apex shape ( Table 1 View TABLE 1 ); besides, the type locality of R. strigosa setosa is within the range of localities where we found these populations ( Fig. 11 View FIGURE 11 ). However there are other characteristics such as the presence of testicular sacs and the lack of lateral typhlosoles that separate R. strigosa setosa from the populations here studied. Furthermore, some of the studied individuals have characters exclusively found in the Guatemalan population of R. strigosa strigosa , like lateral typhlosoles and transverse rows of circular areas in the genital marks (Chajul and Huimanguillo). For the time being, we will consider all specimens found in our survey as R. strigosa setosa . Examination of the types of both R. strigosa strigosa and R. strigosa setosa , sampling in more localities of southern Mexico and the highlands of Guatemala, and the use of DNA sequences (e.g. COI and other genes) will enable robust phylogenetic analyses and clarify whether (or not) i) R. strigosa strigosa is synonymous with R. eiseni , ii) R. strigosa setosa should be elevated to species rank, and iii) whether there are more than one species in the complex of R. strigosa setosa .
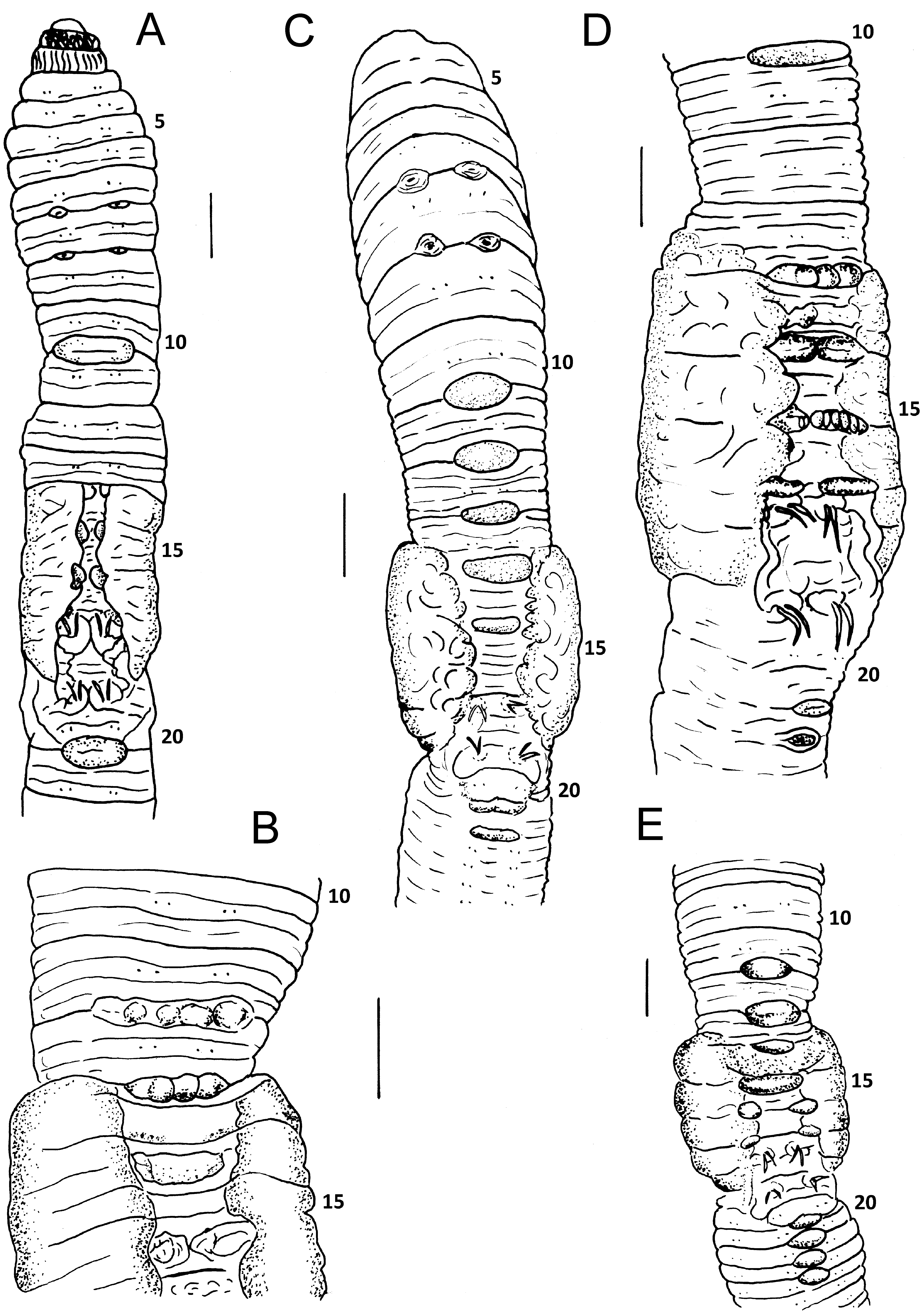
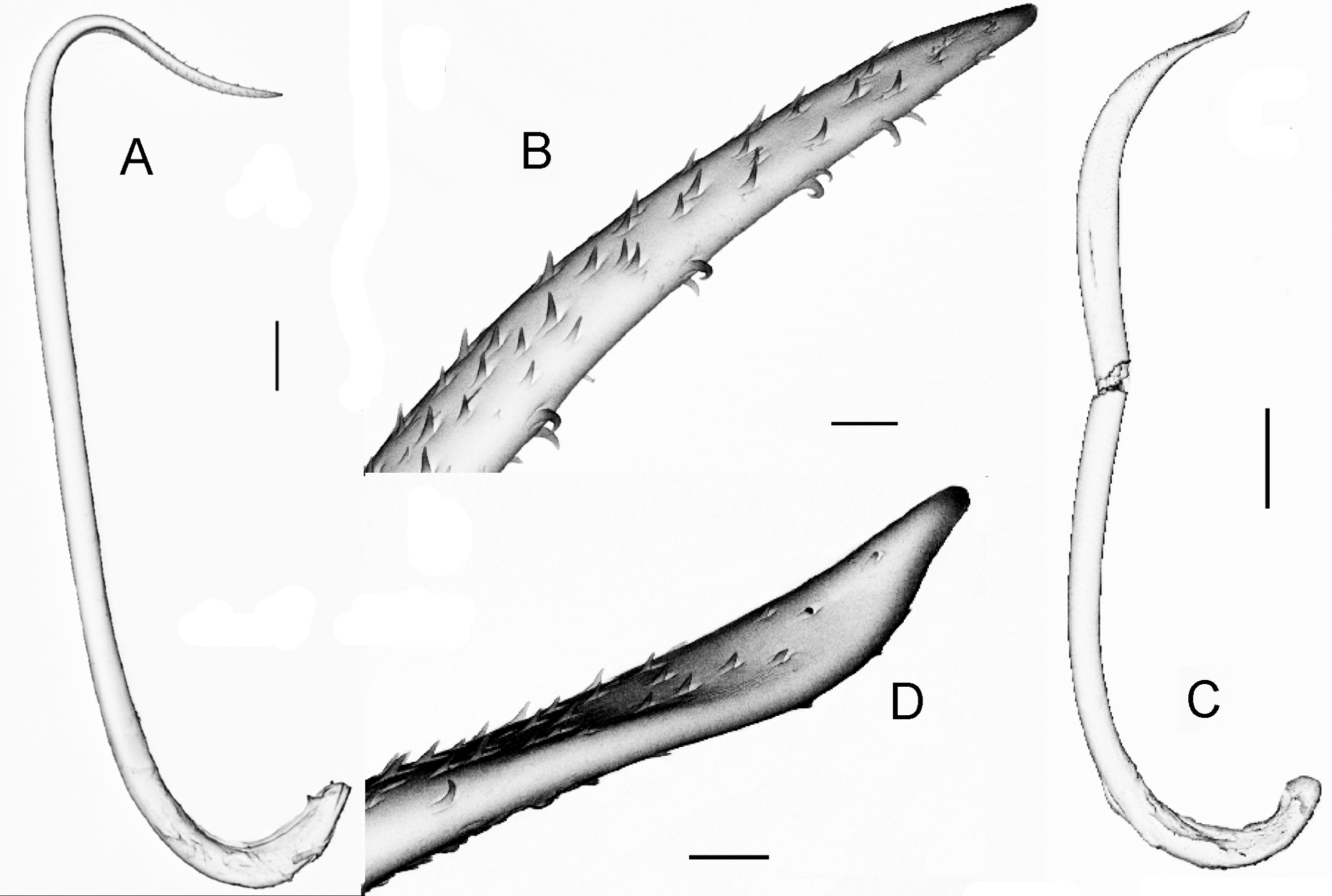
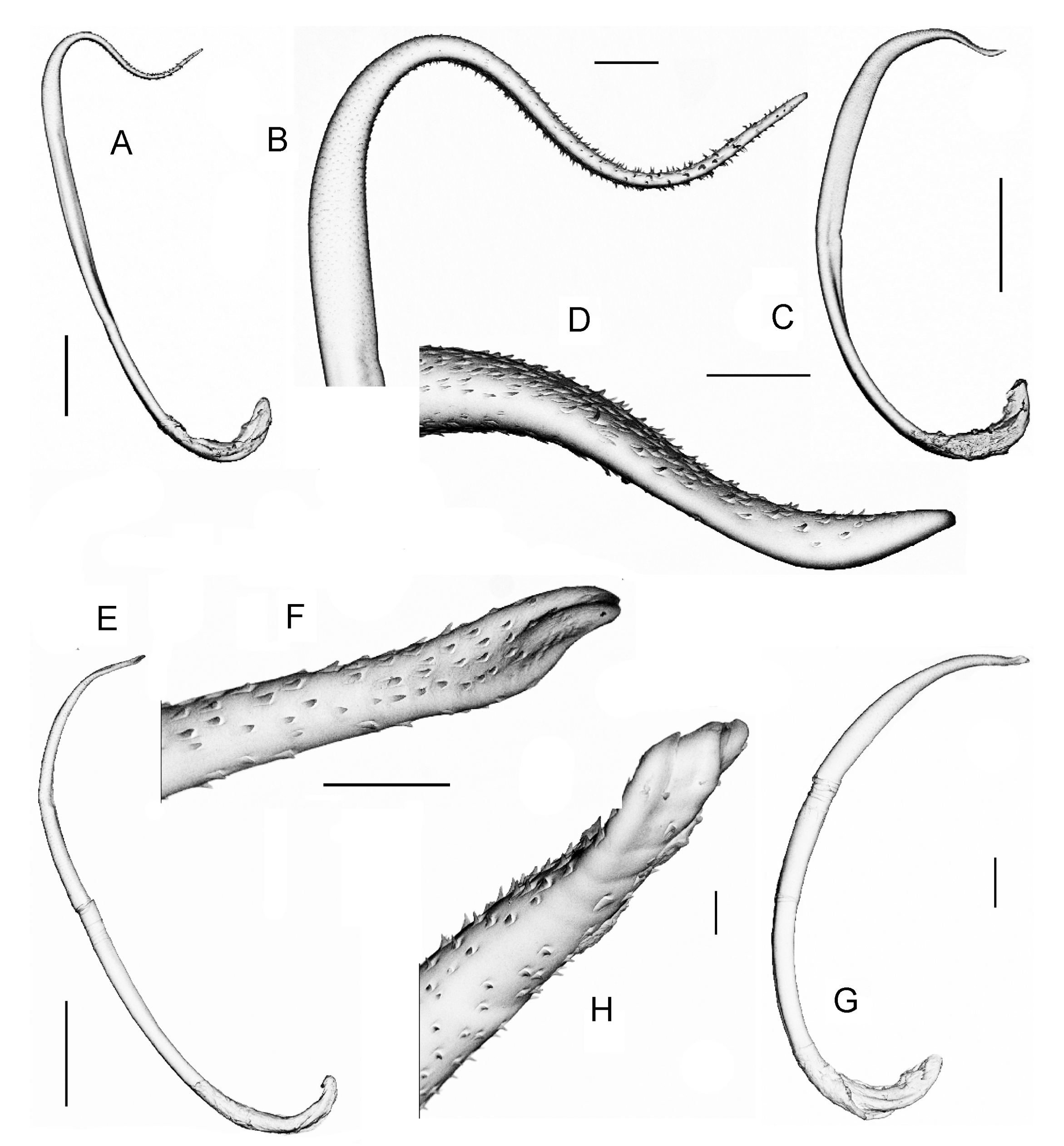
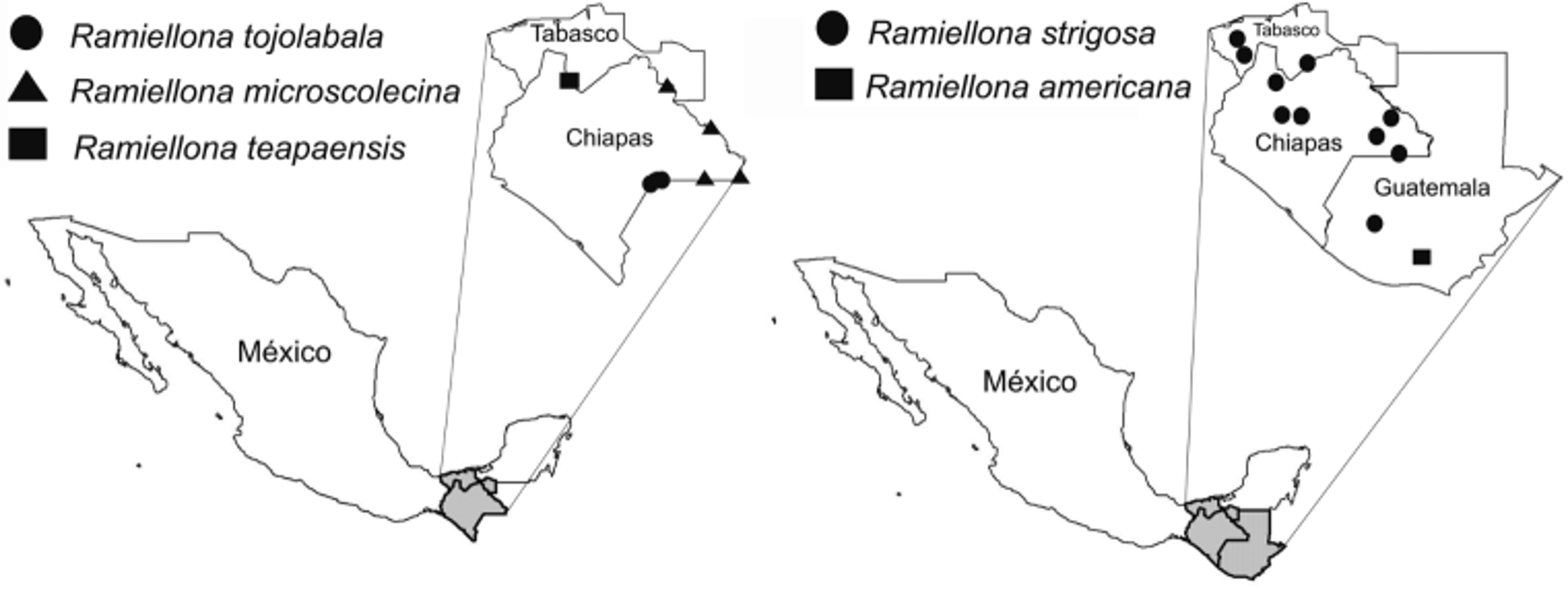
TABLE 1. Morphological comparisons between Ramiellona strigosa setosa Righi from Mexico (this study, last column)
| Character | R. strigosa strigosa Totonicapan (GUA) | R. eiseni Huehuetenango (GUA) | R. strigosa setosa R. strigosa setosa Chiapas (MEX) Chiapas and Tabasco (MEX) |
|---|---|---|---|
| Length (mm) | 97 | 124 | 190 80–200 |
| Width (mm) | 3–3.5 | 3–4 | 7 3.6–6.1 |
| Segment number | 181–208 | ca. 220 | 254 206–328 |
| First setae | In 2 | In 6 (ab), 9 (cd) | In 3 In 3 or 4 |
| Setae before clitellum Setae after clitellum | ab = cd, aa <bc aa> bc | ab = cd, 3.4 aa = bc ab = cd, aa = bc | Not indicated ab = cd, aa<bc ab= 1.5 cd, aa =1.3 bc ab = cd, aa = bc |
| Setae in caudal region | aa and bc ≈ ab and cd | 1.5 ab = cd, 1.3 aa = bc | 1.4 ab = cd, aa = bc ab <cd, aa ≈ bc |
| 1St dorsal pore | 12/13 | 12/13 (11/12 rudimentary) | not recorded 12/13 |
| Clitellum | 13–20 (8) | 1/n 13 –1/n 19 (7) | 14–18 (5) 13, 14–18, 19, 20 (5-8) |
| Genital marks | Unpaired in 10/11, 11/12, 20/21; with transverse rows of circular areas. | 9 unpaired in 10/11, 11/ 12, 14/15, 15/16 and 20/ 21–25/26; paired in 16; those of 15/16 almost paired. | 4 unpaired in 10/11, 14/ Unpaired, 2–9 marks in 15, 19/20 and 20/21; 9/10–15/16, 19/20–24/ paired in 15/16 and 16/17 25; some with transverse rows of circular areas; paired 0–5 marks, in 14/ 15–18/19 |
| Calciferous lamellae | In 8–12 | Absent; esophagus swellings in 12, 13 | In 8–12 In 8–12 |
| Typhlosole | Trifid from 24, with lateral vertical ridges | Trifid from 24, with lateral vertical ridges | Trifid from 21; lateral Trifid from 22,23; with vertical ridges not lateral vertical ridges mentioned |
| Lateral typhlosoles | In 23–28 | Not mentioned, absent? | Not mentioned In the region of 21–29 |
| Caeca | Two in 23–24 or five in 24–28 | Apparently absent | Three in 22–24 Three-four in 22–27 |
| Tufted meronephridia | 3/4 | Not mentioned | Not mentioned In the region of segments 2–5 |
| Septal meronephridia | 4/5–11/12, 12/13 | Not mentioned | Not mentioned Yes, in preclitellar segments |
| Parietal meronephridia | 6 (3/side), from 13 | 8–10 (4–5/side) | 10 (5/side) from 14 8–16 (4–8/side), from 14 |
| Testes | Holandric | Holandric | Holandric Holandric |
| Testicular sacs | Absent | Absent | Present Absent; abundant coagulum |
| Seminal vesicles | Two pairs in 9,11,12 | One pair in 12 | Two pairs in 9 <12 Two pairs in 9 <12 |
| Spermathecae | Two pairs, 2–3 mm length | Two pairs | Two pairs, 5 mm length Two pairs, 4.4–5.7 mm length |
| Ampulla | In 8 and 9, narrowed entally | In 8 and 9, narrowed entally | In 8 and 9, not narrowed In 8 and 9, sausage like entally |
| Duct | Shorter than ampulla | Shorter and wider than ampulla | Thick, shorter than Shorter and wider than ampulla ampulla |
| Diverticle | In 7 and 8, circular flattened disc with small seminal chambers | In 7 and 8, circular flattened disc with small seminal chambers | In 7 and 8, circular In 7 and 8, circular flattened disc with small flattened disc with small seminal chambers seminal chambers |
| Penial setae | 1.5 mm length, curved like parentheses | 2 mm length, curved | 2.5 mm (a) length, 1–2.6 mm length, curved curved like parentheses |
| Dimorphic penial setae | No | No | Yes Yes |
| Apex | Thin and curved | Thin and curved, claw like | a = thin; b = spoon-like a = thin; b = spoon like |
| Ornamentation | Two longitudinal rows of transverse serrations of 2–7 thorns | Absent in apex; last distal third with several transversal teeth | Limited to apex, with Last distal third, several irregular thorns including apex, with several irregular thorns |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ramiellona strigosa setosa Righi, 1972
| Fragoso, Carlos & Rojas, Patricia 2014 |
Ramiellona strigosa setosa
| Righi 1972: 222 |