
Heteropsis tornado, C, Lees David, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4118.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:CFA586DA-10EE-468B-80EE-35351E3845FD |
|
DOI |
https://doi.org/10.5281/zenodo.6086430 |
|
persistent identifier |
https://treatment.plazi.org/id/03874732-4C5A-C67A-1EB7-29EDFCB32348 |
|
treatment provided by |
Plazi |
|
scientific name |
Heteropsis tornado |
| status |
|
Heteropsis subsimilis group
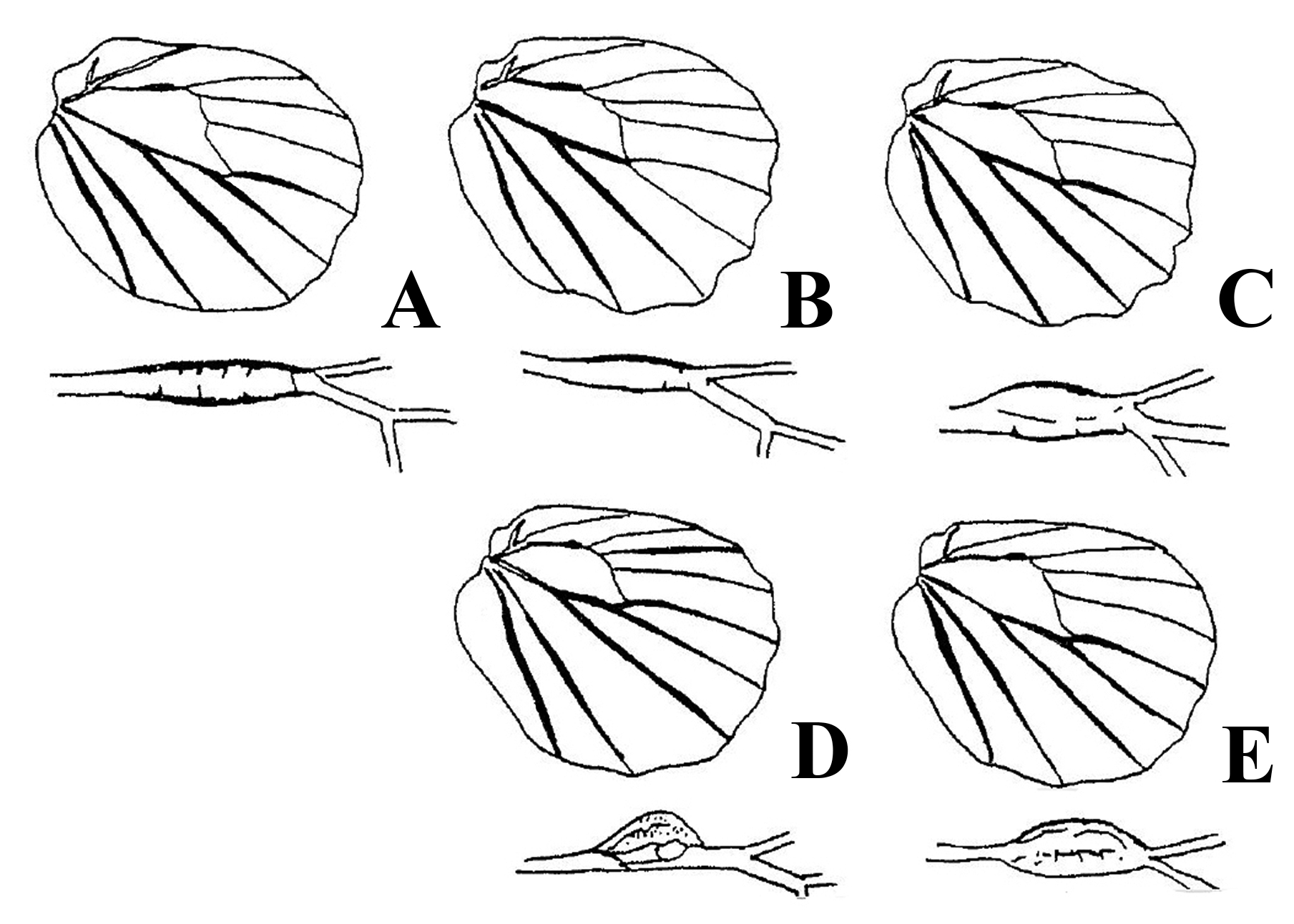
The ‘ subsimilis 7’ species group (after Pseudonympha subsimilis Butler, 1878 ) of Torres et al., (2001) resemble each other not only in wing pattern, but also in ♂ genitalia. This Ht. subsimilis group, which occurs within Malagasy rainforest, has also colonised the Comoros, as H. comorana ( Oberthür, 1916) , a species closely related to the generally abundant Ht. pauper ( Oberthür, 1916) ( Lees, 1997: 168; Torres et al., 2001). The sister grouping of the Ht. subsimilis group was unresolved in several earlier studies ( Lees, 1997, Torres et al., 2001, Linares et al., 2009, Kodandaramaiah et al., 2010), although these studies found good support that the Ht. subsimilis species group forms a clade (97% based on COII parsimony analysis: Torres et al., 2001; 100% based on maximum likelihood analysis of COI, Ef1a and wingless: Kodandaramaiah et al., 2010). Linares et al., (2009) presented a supported phylogeography of relationships of three species. Recently though, Aduse-Poku et al., (2015, Fig. 1 View FIGURE 1 ) found strong support for three sampled species namely Ht. subsimilis , Ht. pauper and Ht. comorana , which clade they recovered as sister to 13 sampled species of the Ht. strigula group, including Ht. tornado sp. nov. and the Ht. ankova subgroup (see below). ♂ genitalia have valve tips with a stout inward projecting spine that fall far short of the uncus extension; from LV, the uncus is inflated before its tip. The ♂ genitalia are in fact remarkably miniaturised compared to other Heteropsis , only around 1.7 mm in length (apart from those of Ht. tornado which is about 2 mm long, it can be seen in figures here that those of the Ht. strigula group are generally between 2–3 mm long and the Ht. drepana and Ht. antahala groups are more than three mm long and exhibit extension of particular parts). It may be hypothesised that such reduction comes with a relaxation of importance of this character system as a pre-mating isolation mechanism, as apparent in the Ht. subsimilis group, yet only in the Ht. drepana group is there a tendency to reduction or even loss (the supra-discocellular patch of Ht. narova ) of parts of the androconial system (perhaps regained in Ht. drepana only). Atrophication of the genitalic system may be compensated for in a different, possibly androconial character system. Phenotypically, adult ♂♂ and ♀♀ of the Ht. subsimilis group are similar, nearly monomorphic except in size, but for simple androconia in the ♂ discocellular region and an inflated HW vein system also found in the Ht. strigula group. In the Ht. subsimilis group, this system features a set of linearly rather than bulbously inflated veins in the ♂ HW ( Fig. 12 View FIGURE 12 ; Lees, 1997: 96). As predominant in the Ht. strigula group, all species have the reduced ocellus configuration of the HWD, with only that in space CuA1 expressed as well as often that in space Rs; the morphological ‘total evidence’ phylogenetic hypotheses of Lees (1997: 156) that the Ht. subsimilis group is sister to only part of the Ht. strigula group (see also Torres et al., 2001) is presumably biased by the inclusion of this character system, whereas the unsupported Bayesian topology in Kodandaramaiah et al., (2010, Fig. 3 View FIGURE 3 ) would imply that this configuration of HWD reduction is convergent between these two sister species groups (only with a gain of expression in HWD space-M 3 in Ht. roussettae sp. nov.). In any case, the two groups are sisters ( Aduse-Poku et al., 2016, in press). Both sexes are attracted to ripe fruit as adults, as well as sometimes other sugar sources, and these low flying butterflies seem to specialise on grasses in the forest interior. There seem to be ridge and riparian specialists ( Kremen, 1994).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Satyrinae |
|
Tribe |
Satyrini |
|
Genus |