
Pseudocandona lordi, Külköylüoğlu & Hutchins & Yavuzatmaca & Schwartz, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5046.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:C0B54B87-57A9-456D-8942-D16EEB1678B5 |
|
persistent identifier |
https://treatment.plazi.org/id/F627B379-FF87-5E6A-FF6F-FC1AFCCC138C |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudocandona lordi |
| status |
sp. nov. |
Pseudocandona lordi n. sp.
( Figs 13–18 View FIGURE 13 View FIGURE 14 View FIGURE 15 View FIGURE 16 View FIGURE 17 View FIGURE 18 )
Type locality. Rio Grande River Basin , Live Oak Creek , Crockett County, Texas, USA (Lat: 30.742 196, Long: - 101.673946), Elevation ~ 656 m ASL .
Etymology. The species is named after Prof. Dr. Alan R. Lord (Senckenberg Forschungsinstitut, Germany) for his contributions in the study of Ostracoda.
Material examined. Holotype. Male dissected in lactophenol solution with soft body parts slide, mounted and sealed with translucent nail polish (no: OK-TX-LOCr-150713 H3-1); valve kept in micropalaeontological slide (no: OK-TX-LOCr-150713 H3-2). Collected from type locality on 13 July 2015 by Benjamin T. Hutchins and Danielle Belleny.
Allotype. One female from the type locality (no: OK-TX-LOCr-150713 H3-3). Collected by Benjamin T. Hutchins and Danielle Belleny.
Dissected paratypes. One male (no: OK-TX-LOCr-150713 H1-4) and two females (no: OK-TX-LOCr-150713 H3-5-6) dissected in lactophenol solution with soft body parts sealed with translucent nail polish. Collected from type locality by Benjamin T. Hutchins and Danielle Belleny.
Non-dissected paratypes. Five females, one male, four juveniles, and two valves kept in 70% ethanol. Collected from type locality by Benjamin T. Hutchins and Danielle Belleny.
Repository. The holotype and all paratypes are stored at the Limnology Laboratory of the Biology Department, Bolu Abant İzzet Baysal University, Bolu, Turkey .
Description: Measurements (in µm). Male (n=3): LV: 1100–1140 (mean = 1120), H: 800–830 (mean =815); RV: L=1040–1100 (mean = 1070), H=800–810(mean = 800); W=590. Female (n=3): LV: 1100–1160 (mean = 1080), H: 800–840 (mean = 820); RV: L=1040–1110 (mean = 1070), H=810–830 (mean = 820); W=600.
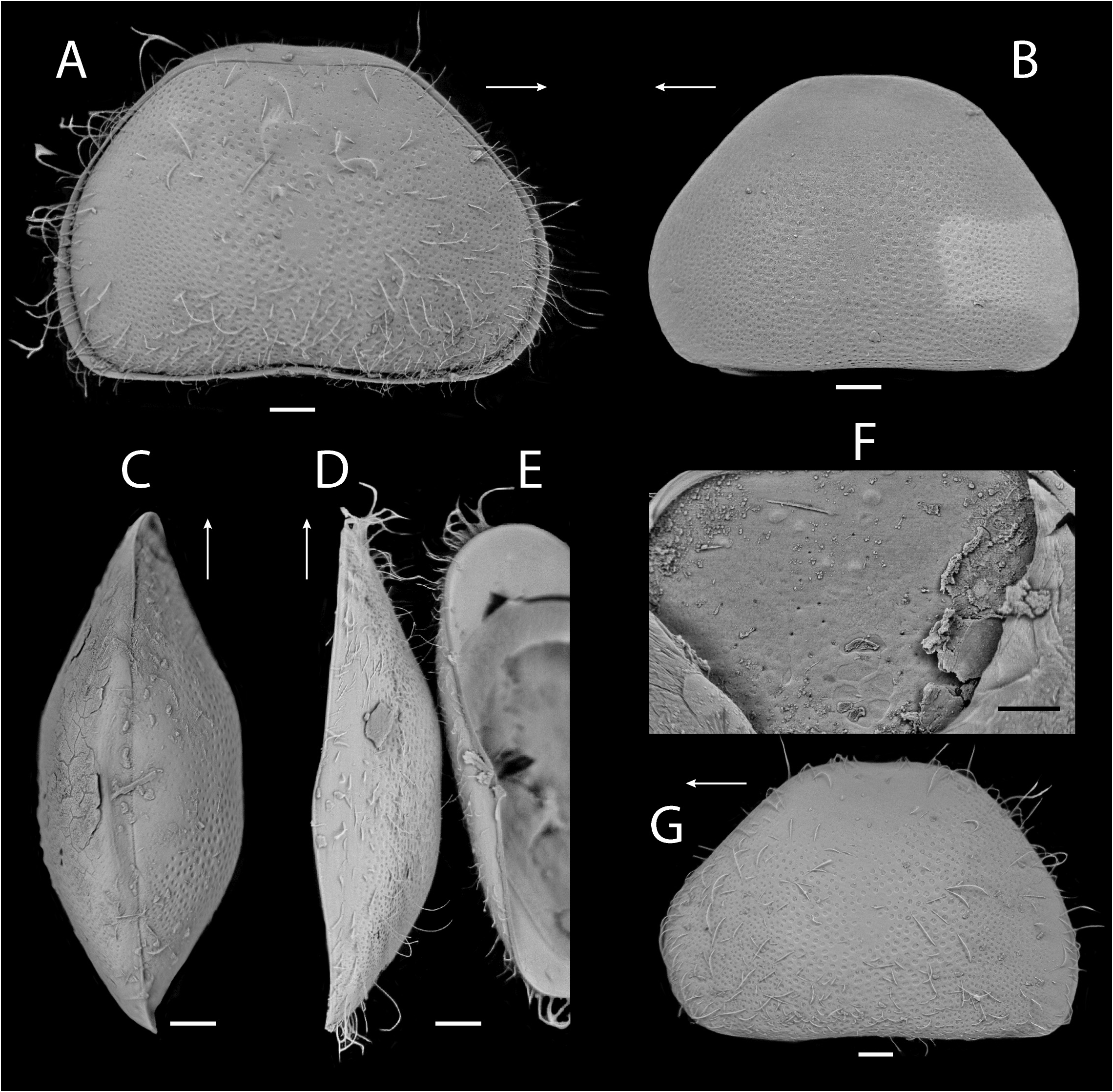
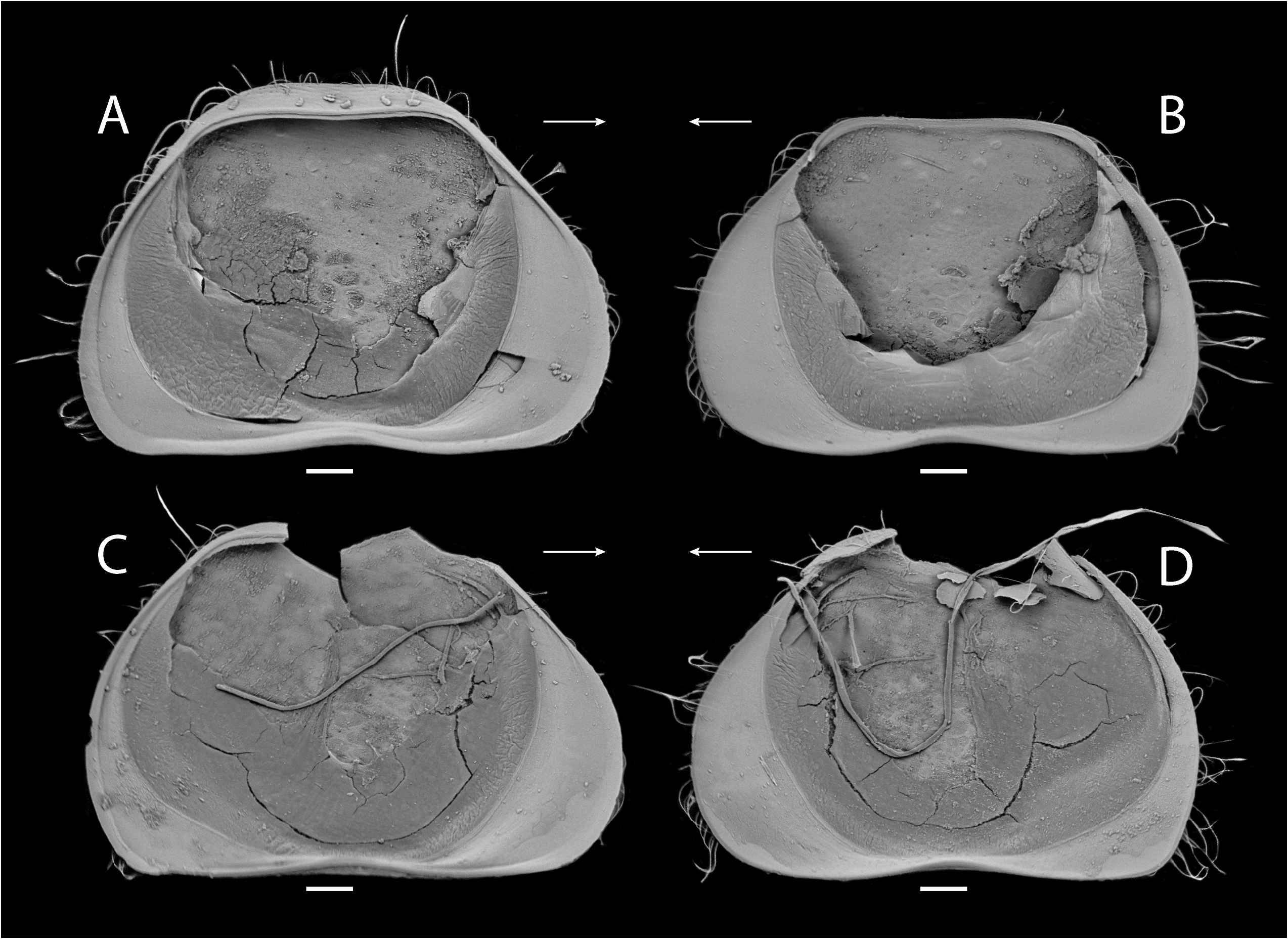
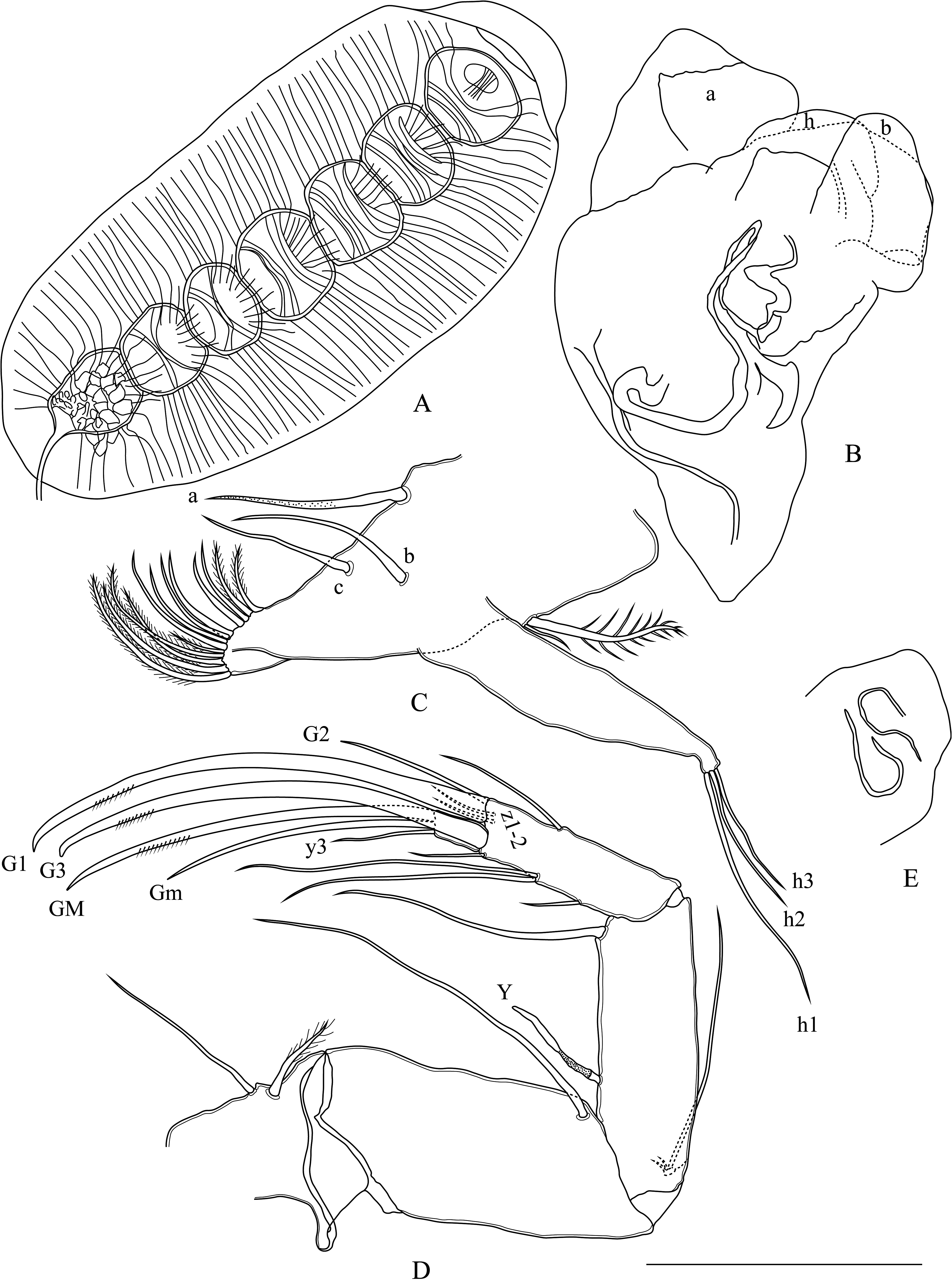
Carapace ( Figs. 13–15 View FIGURE 13 View FIGURE 14 View FIGURE 15 ) opaque to white, suboval in lateral view, surface with small to large pits and long setae ( Figs. 13A, B View FIGURE 13 ), dorsally concave in center (less concave in ventral view) ( Fig. 13C View FIGURE 13 ). Anterior margin rounded ventrally ( Figs. 13D, E View FIGURE 13 ). Posterior margin rounded, ventrally with acute point. Margins without spines or tubercles. Greatest height near center. Carapace thin in dorsal view, navicular shape, with maximum width near center, margins slightly pointed at ends. LV more concave than RV dorsally. LV protruding over RV at center in ventral view. In internal view ( Fig. 13F View FIGURE 13 ), rounded tubercles present dorsally, calcified inner lamella wider anteriorly than posteriorly. LV overlapping RV on all margins ( Fig. 13G View FIGURE 13 ). Uncalcified part wide ( Figs. 14A–D View FIGURE 14 ). Inner list and groove absent. Five central muscle scars present.
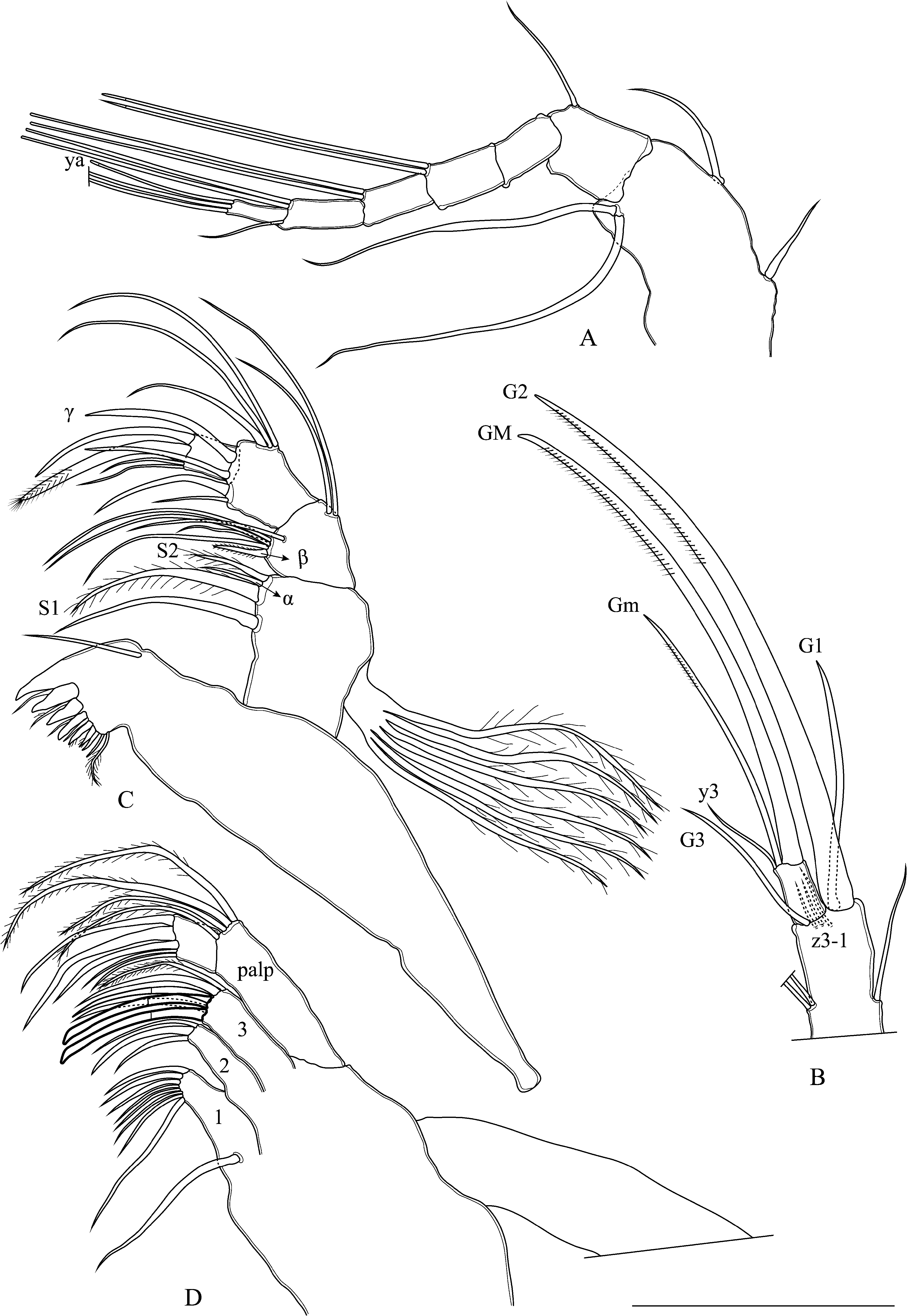
A1 ( Fig. 16A View FIGURE 16 ): Seven-segmented. Wouters and Rome organs absent. First segment with two smooth ventral setae reaching end of penultimate segment and with medium size smooth dorso-median seta. Second segment with thin seta dorso-anteriorly. Third segment without seta. Fourth segment with one claw-like and one smooth seta.. Fifth segment with two claw-like setae. Penultimate segment with one short ventral seta and two claw-like long setae. Terminal segment with two very long claw-like setae. Aesthetasc ya twice as long as terminal segment.
A2 ( Fig. 16B View FIGURE 16 ): Four-segmented (distal parts drawn). First segment with one long smooth basal seta. Exopodial plate with two very short setae and one long thin seta about as long as second segment. Second segment with aesthetasc Y about 1/3 as long as second segment and one long smooth posterior seta. Natatory setae absent. Penultimate segment undivided, y1 and y2 aesthetasc short, slightly longer than terminal segment, with two long smooth t-setae, z setae (z1–3) very short about size of terminal segment, G1 claw thin, about 1/3 as long as G2, G2 claw long and well-developed with pointed tip, G3 claw very short, seta-like, twice as long as terminal segment, aesthetasc y3 short, seta-like. Terminal segment with a well-developed GM claw, Gm claw thin, about half as long as GM claw.
Md-palp ( Fig. 16C View FIGURE 16 ): Md-coxa well-developed, with 6 strong teeth. First segment with two plumose S-setae (S1 twice as long as S2) and one smooth seta about as long as S1, alpha seta short and thin. Vibratory plate with six plumose setae. Second segment with group of four unequally long setal group (+beta seta stout and 1 slightly hirsute Penultimate segment with three unequally long smooth dorsal setae, gamma seta smooth (twice as long as terminal segment), with three apical setae. Terminal segment rectangular in shape, with two claw-like (second slightly plumose) and two thin setae.
Mx1 ( Fig. 16D View FIGURE 16 ) with three endites and Mx1-palp. First endite with seven short and one long setae. Second endite with two smooth bristles and two smooth seta. Third endite with two short (apical seta plumose), two medium setae and two smooth bristles (second one slightly longer). Vibratory plate with 21 long setae. First palp with four unequally long plumose setae. Terminal palp subrectangular, with two short and three well-developed claw-like smooth setae.
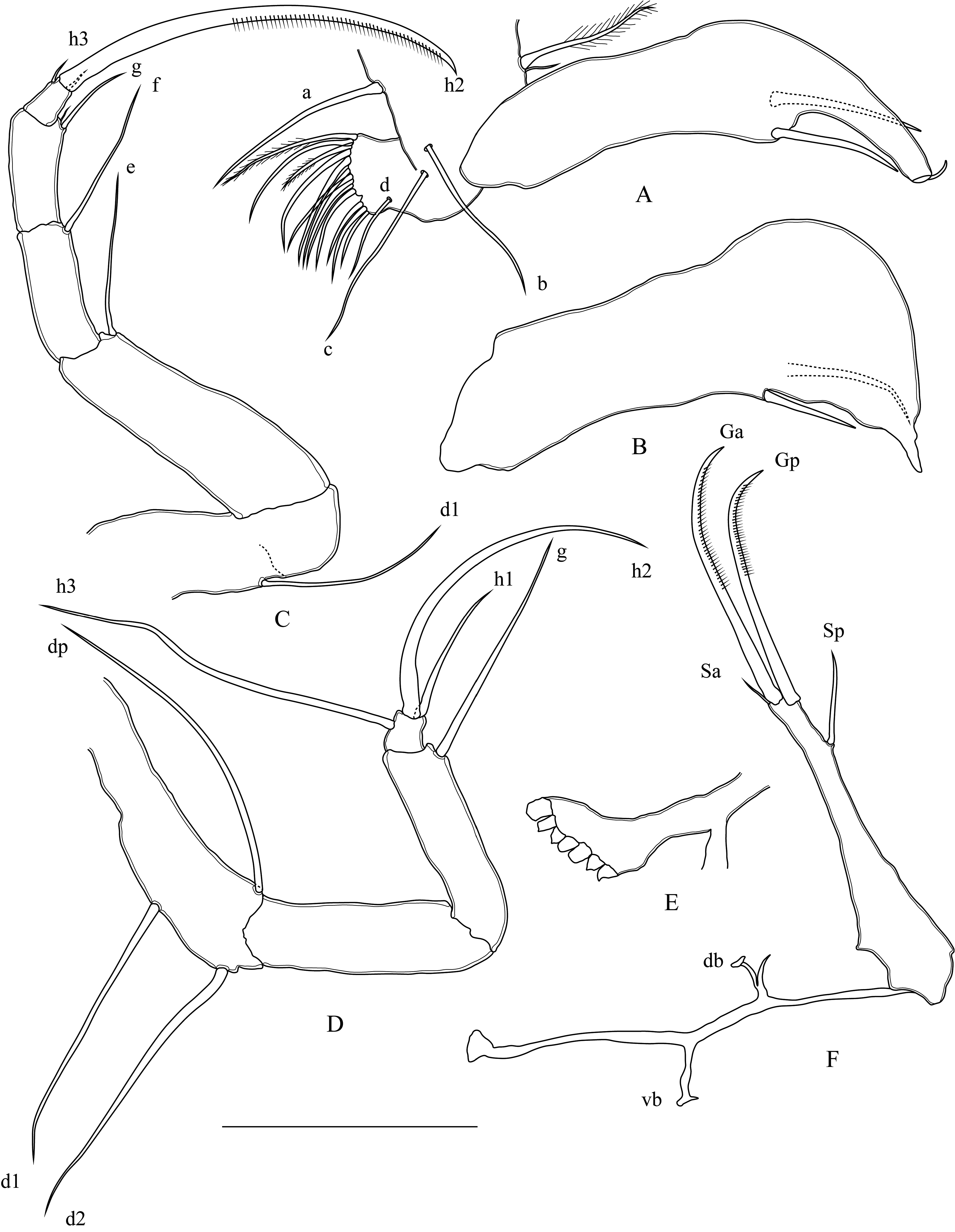
T1 ( Figs.17A, B View FIGURE 17 ): Transformed into prehensile palps. Setae a, b, and c relatively long, seta d not apparent. Vibratory plate with one long plumose and one short setae. Right prehensile palp ventrally enlarged, with pointed end, with two well-developed apical spines, left palp longer, ending with a hook like finger.
T2 ( Fig. 17C View FIGURE 17 ): Five-segmented. First segment with one long and smooth d1 seta, setae dp and d2 absent. Setae e, f, and g smooth, length ratio: e ≈ f> g. Terminal segment subrectangular in shape, with two short (h1, h3) setae, and h2 seta long and claw-like.
T3 ( Fig. 17D View FIGURE 17 ): Four-segmented. d1, d2, and dp setae long. Setae e and f absent, g seta long (about 5x as long as terminal segment). Terminal segment square in shape, with two long and one short setae.
Rake-like organ ( Fig. 17E View FIGURE 17 ): Ventrally branched, with seven short teeth.
Uropod ( Fig. 17F View FIGURE 17 ): with a short ramus, claws well-developed and serrated. Anterior seta short (about 2x as long as posterior claw). Caudal attachment with two ventral branches.
Zenker’s organ ( Fig. 18A View FIGURE 18 ): seven rows (5 + 2) of spines.
Hemipenis ( Fig. 18B View FIGURE 18 ): Outer lobe large and subtriangular, inner and medial lobes relatively rounded.
Female description: Carapace similar to male.T1 with three unequally long and smooth apical setae (h1>h2>h3) ( Fig. 18C View FIGURE 18 ). A2 with short seta-like G2 claw about 1/3 as long as G1 claw ( Fig. 18D View FIGURE 18 ). Other parts of A2 similar to male. Genital organ simple, rounded ( Fig. 18E View FIGURE 18 ). All other soft body parts similar to male.
Accompanying ostracod species. See Appendix 2.
Differential diagnosis. Carapace surface with pits and long setae, dorsally concave. A1 without Wouters and Rome organs, penultimate and terminal segments with claws. A2 without swimming setae and with very short z-setae. T1 vibratory plate with two unequally long setae, clasping organs asymmetrical in male. T2 five-segmented, h2 claw very long. T3 four-segmented with two long and one short setae. Setae e and f absent in T2. Second segment of Md palp with five unequally long setae (+ 1 beta seta) ventrally. Mx1 with two smooth bristles. Hemipenis with well- developed lobes. Zenker’s organ with five medial and two apical rows of spines.
Remarks: Candonidae is one of the most speciose ostracod families and has a widespread geographical distribution. The new species is placed in Pseudocandona due to several shared characteristics (see diagnosis of the genus above) (e.g., see Meisch 2000). However, placement of several species and/or groups of species within Pseudocandona versus Typhlocypris has been an ongoing discussion (e.g., see Karanovic 2005, 2012; Namiotko et al. 2014). The two genera can be distinguished by the right valves which are more or less rectangular in Pseudocandona but triangular in juveniles (A-3, e.g., see T. eremita sensu lato) to quasi-rectangular in adults in Typhlocypris (see details in Namiotko et al. 2014). Pseudocandona lordi n. sp. exhibits the more or less rectangular right valve ( Fig. 15 View FIGURE 15 ) characteristic of Pseudocandona . The genus has four species groups: rostrata, compressa, carribeana and prespica whereas Typhlocypris includes the eremita species group. Pseudocandona lordi n. sp. is assigned to the compressa group because of five setae in the second segment of Md-palp and relatively long h1 setae (ca. three times longer than terminal segment, not curved) of the terminal segment of T3 (see Meisch 2000).
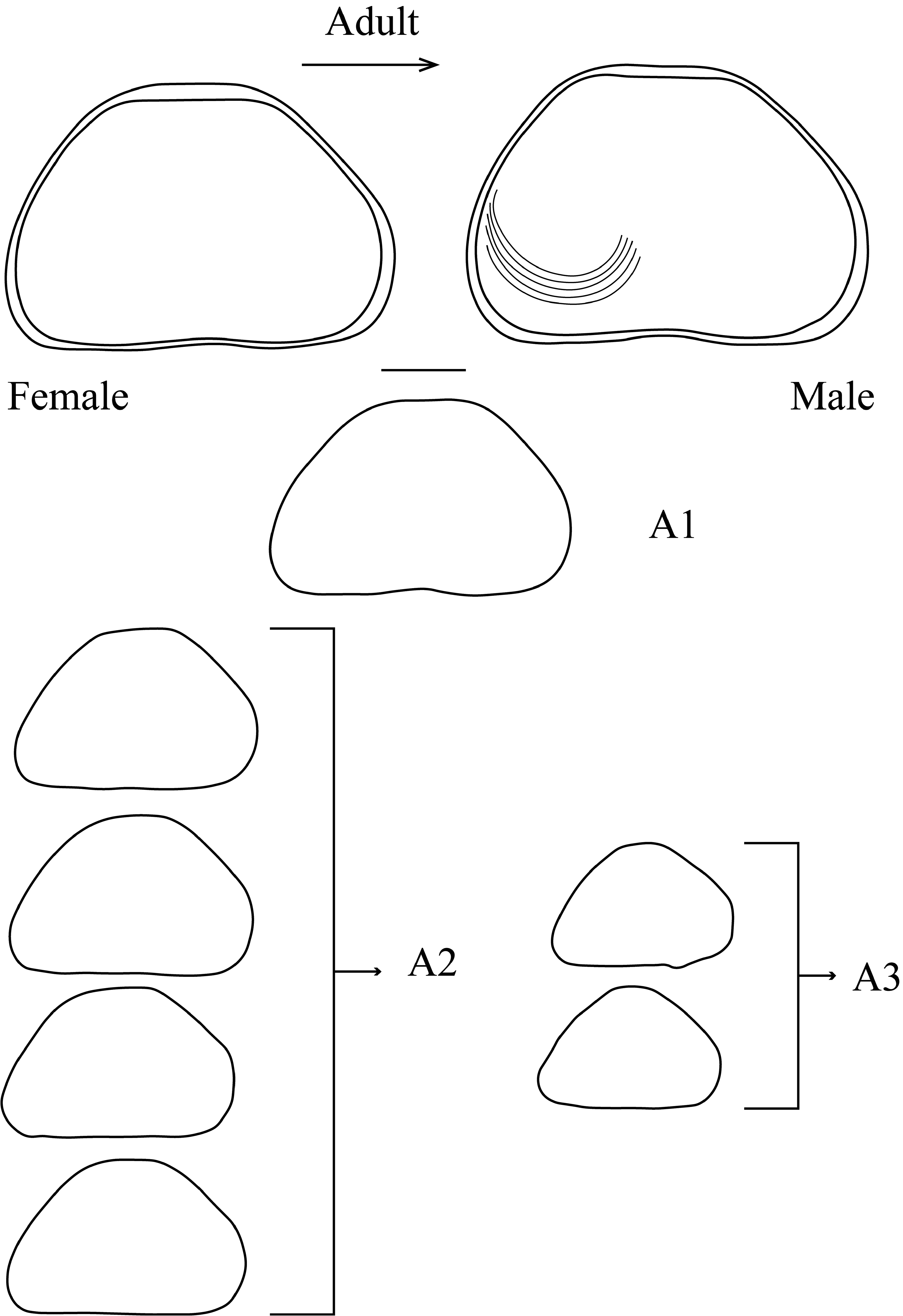
Pseudocandona includes 72 species ( Meisch et al. 2019) with a diversity of unique, habitat-specific adaptations to a variety of aquatic environments (see Discussion). A group of species that includes P. dorsoconcava (Bronstein) bears one claw on the terminal segment and two claws on the penultimate segments of A1 ( Bronstein 1947). Bronstein (1947) suggested that this feature was common in all Lake Baikal species (e.g., Candona sensibilis Bronstein , C. directa Bronstein , C. unguiculata Bronstein ) where it was considered a unique adaptation for digging in sediment on the lake bottom. Similar structures suggest that P. lordi n. sp. may also dig in sediment, except that claws are present on the fourth segment rather than the terminal segments. Pseudocandona dorsoconcava and P. lordi n. sp. are distinguished from one another by several differences in shape and soft body parts. For example, z-setae are transformed into claws in males of P. dorsoconcava but not in P. lordi n. sp., and a well-developed protuberance is present on the ventro-anterior margin of the valves in P. dorsoconcava but absent in P. lordi n. sp.. However, P. lordi n. sp. carapace shape changes during ontogenetic development ( Fig. 15 View FIGURE 15 ). During the first ontogenetic stages of the instars (A1–A3), carapace shape appears more triangular than in adults. Starting with the A3 instar, the protuberance on the LV becomes more visible and the shape becomes more oval. In all stages, LV>RV. Morphological resemblance of P. lordi n. sp. and P. dorsoconcava may be due to several factors such as similarities in habitat preferences. The lack of swimming setae on A2 (a diagnostic characteristics of the subfamily) clearly indicates that both species are bottom-dependent. Additionally, both species bear relatively long terminal claws on A2 and stiff claw-like setae on A1. Bronstein (1947) considered these characters as primitive in P. dorsoconcava and closely associated with Tertiary freshwater species in which they were an adaptation for digging.
Although the evolution of some Texas aquifers may date back to the Cambrian Period ( Ashworth & Hopkins 1995), P. lordi n. sp. (and the other five species described herein) is associated with hyporheic habitats overlying the Edwards and Edwards-Trinity Aquifers, which are of Cretaceous age ( Ashworth & Hopkins 1995) or younger. While the age of Lake Baikal is the subject of debate, the lake probably existed by the Late Cretaceous, but the majority of Lake Baikal species appear to be 3–4 million years old (see details in Mats et al. 2011) or even older. Ostracods are known to be highly diverse ( Karanovic & Sitnikova 2017) that some species flock (e.g., Cytherissa spp. ) may have originated in Lake Baikal ( Mazepova 1990). Several other ostracod species (e.g., Scottia pseudobrowniana Kempf ) have a Holarctic distribution supporting geographical correlation between Nearctic and Palearctic regions ( Külköylüoğlu & Vinyard 2000). However, insight about the zoogeography of P. lordi n. sp. is limited by the lack of fossil record for the species and no knowledge of its distribution beyond the type locality. Further studies across the Holarctic will be needed to interpret the morphological resemblance between P. dorsoconcava and P. lordi n. sp.
Ecology: Pseudocandona dorsoconcava was reported from Lake Baikal at depths of 4.5 m (17.5°C) to 75 m (3.82°C) ( Bronstein 1947). Pseudocandona lordi n. sp. was collected from approximately 0.5 m depth in the hyporheic zone of Live Oak Creek (see Appendices 2, 3). Although subsurface flow was probably occurring within the hyporheic zone, surface water was reduced to a few stagnant pools on the date of collection. Hyporheic water in which P. lordi n. sp. was collected had the following physical parameters: water temperature 24.6°C, dissolved oxygen 4.57 mg /L, specific conductance 2110 µS/cm, and pH 7.35. Pseudocandona lordi n. sp. occurs with a diverse assemblage of stygobionts that include Erimidrilus annelids, the mite Arrenurus , the amphipods Parabogidiella cf. americana (Holsinger) , Seborgia hershleri (Holsinger) , and Artesia subterranea (Holsinger) , the bathynellid Tex- anobathynella, and the isopod Lirceolus cf. cocytus (Lewis) .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.