
Psammoleptomesochra Mielke, 1994a
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5051.1.13 |
|
publication LSID |
lsid:zoobank.org:pub:F94203E7-FCD1-4975-BAD3-0DF534806712 |
|
DOI |
https://doi.org/10.5281/zenodo.5576521 |
|
persistent identifier |
https://treatment.plazi.org/id/951887EA-FFFB-FFA7-FF51-D173E16AFCF4 |
|
treatment provided by |
Plazi |
|
scientific name |
Psammoleptomesochra Mielke, 1994a |
| status |
|
Psammoleptomesochra Mielke, 1994a
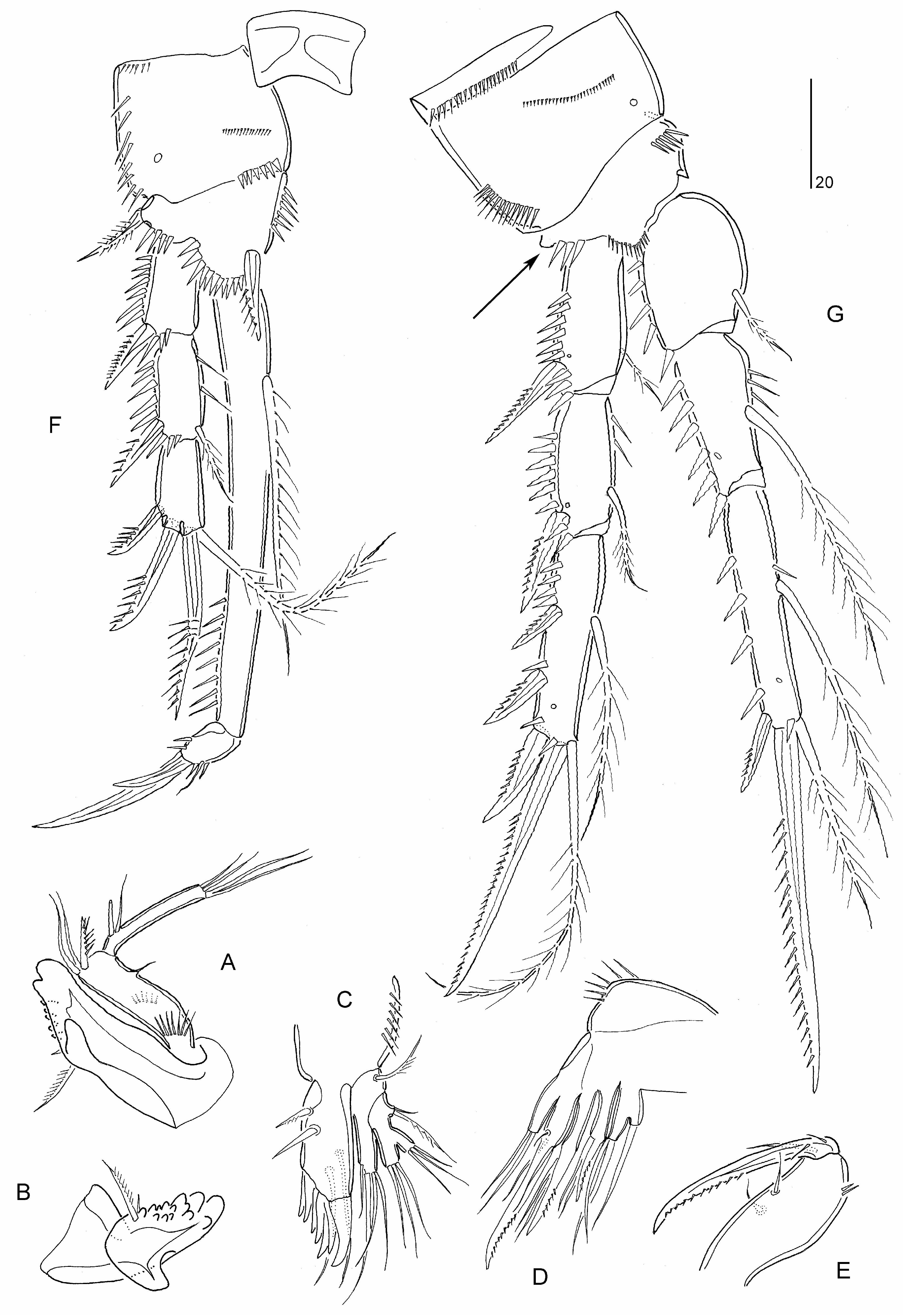
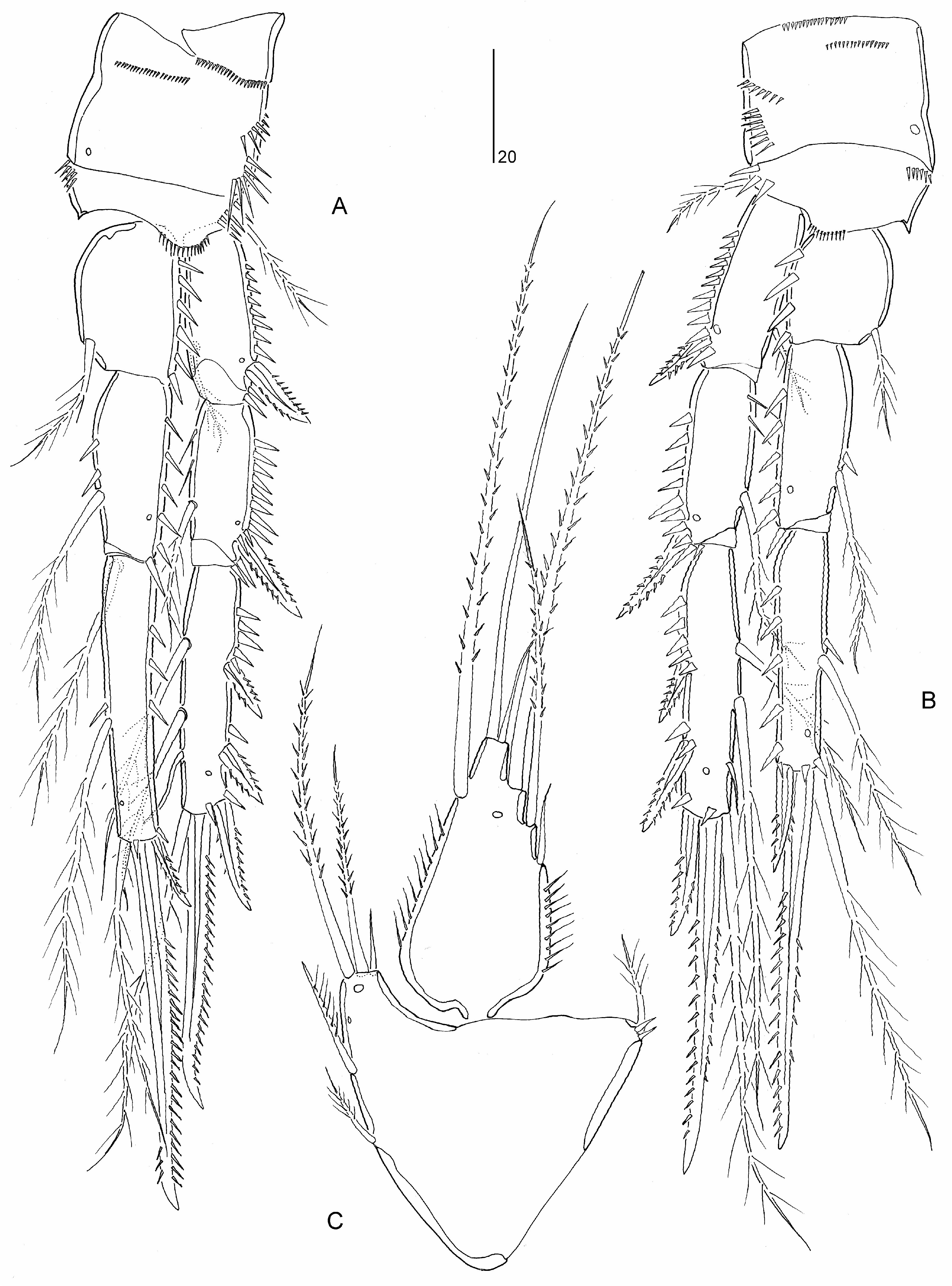
Mielke (1994a: 258) placed Psammoleptomesochra in the “ Leptomesochra -complex”, an artificial group of genera in the Ameiridae ( Lang 1965; Wells 1967), and recognized a certain closeness with Parapseudoleptomesochra reducta Rao, 1972 (now type species of Raoleptomesochra Karanovic, 2004 ) and particularly Paraleptomesochra . Psammoleptomesochra displays many autapomorphic character states including, (a) antennary exopod without lateral setae on exp-2; (b) mandibular basis with two setae; (c) mandibular exopod vestigial, represented by minute seta; (d) coxal epipodite on maxillule absent; (e) maxillipedal basis without setae; (f) P1 exp-2 without inner seta; (g) P2 exp-3 without inner setae; and (h) P2–P4 enp-3 with reduced distal setae (represented by two setules).
Psammoleptomesochra shares the area of reduced chitinization on the elongate P1 enp-1 with Paraleptomesochra and Foweya , while the sexual dimorphism of the inner basal spine on P1 is also displayed in Foweya and Thalestrella . Paraleptomesochra has many reductions in common with Psammoleptomesochra including the loss of setation elements on the mandibular basis and exopod, maxillipedal syncoxa and basis, P2–P4 and endopodal lobe of ♀ P5 ( Table 2 View TABLE 2 ). In both genera the armature of P2–P4 is strongly reduced but not all setal losses are evidence for their common ancestry. For example, as a result of convergent evolution, both genera display only four elements on P3 exp-3 but in the former the pattern consists of one inner, two distal and one outer element(s) [121] while in the latter there are two distal and two outer elements [022].
Diagnosis. Parastenheliidae . Sexual dimorphism in antennule, P5–P6 and urosomal segmentation. Body subcylindrical; posterior margin of cephalothorax and all somites (except anal somite) with deeply divided, denticulodigitate, hyaline frills. Rostrum defined at base, small, reaching to proximal third of second segment of antennule; linguiform with rounded apex. Anal operculum semicircular, bordered with few (around six) strong spinules. Caudal ramus about twice as long as wide, with dorsal oblique row of spinules; with six setae, setae II–III short, pinnate and spiniform, setae IV–V not swollen at base.
Antennule short and 9-segmented in ♀, segment 1 not elongate, segments 7–8 shortest, with aesthetascs on segments 4 and 9; haplocer in ♂, segmentation, position of geniculation and segmental homologies unconfirmed. Antenna with basis; proximal endopodal segment with short abexopodal seta; distal endopodal segment without penicillate elements; exopod 2-segmented, proximal segment with two lateral setae, distal segment with two apical elements. Mandible with two elements on basis; endopod with two lateral and 5–6 apical setae; exopod vestigial, represented by short seta. Maxillulary coxal epipodite absent. Maxilla with three endites on syncoxa; endopod discrete, with three setae. Maxilliped with one seta on syncoxa; basis without seta(e) but with short longitudinal spinular row on palmar margin; endopod represented by claw with one accessory seta.
P1 inner basal spine sexually dimorphic, recurved in ♂. P1 exopod 3-segmented, segments subequal in length; exp-2 without inner seta; exp-3 with two pinnate spines and two geniculate setae. P1 endopod 2-segmented; enp-1 longer than exopod, with well developed, pinnate inner seta arising from area of reduced chitinization in middle third of segment; enp-2 with one naked minute seta and two claws (one geniculate, one non-geniculate). P2–P4 rami 3-segmented; endopods short, at most as long as exp-1 and -2 combined; inner distal element of P2–P4 exp-3 setiform and thin; endopodal segments without inner setae except for P4 enp-1 bearing long pectinate seta; P2–P4 enp-3 with two vestigial distal setae and one outer spine. P2–P4 endopods ♂ without sexual dimorphism. Armature formula of P2–P4 as follows:
P 5 ♀ endopodal lobe with four elements, innermost spiniform; inner margin without transverse striae; exopod elongate, with six naked setae. P 5 ♂ endopodal lobe with two elements, outer one shortest; exopod 1-segmented, with five elements, inner one vestigial. Vestigial P 6 ♀ represented by two setae. P 6 ♂ with two setae and one spine (innermost element).
Type and only species. Psammoleptomesochra australis Mielke, 1994a (by original designation).
Type locality. Chile, Los Lagos Region, Chiloé archipelago, southeastern coast of Isla Chiloé, Quellón Viejo ; sandy beach .
Original description. Mielke (1994a): 252–258; Figs 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 .
Notes. The endopodal armature pattern adopted herein differs from Mielke’s (1994a: 257) formula in two aspects: the apical spiniform element on enp-3 is here considered as the homologue of the outer spine in other genera, and the two minute and fine setae originating from the inner corner of enp-3 are included in the formula as distal elements ( Table 1 View TABLE ). Mielke (1994a) observed variability in the armature of the P5 endopodal lobe (with 2–3 apical setae) in the female and the P5 exopod (distal seta on outer margin variable in length) in the male.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |