
Prionus (Homaesthesis) simplex ( Casey, 1912 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4134.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:92AC0E20-F532-4D21-AE1F-4B056327212F |
|
DOI |
https://doi.org/10.5281/zenodo.5066931 |
|
persistent identifier |
https://treatment.plazi.org/id/03AA87AC-FFD7-671B-FF2C-C1432E1F85B4 |
|
treatment provided by |
Plazi |
|
scientific name |
Prionus (Homaesthesis) simplex ( Casey, 1912 ) |
| status |
|
Prionus (Homaesthesis) simplex ( Casey, 1912) View in CoL
( Figs. 12–13 View FIGURES 1 – 19 , 54–68 View FIGURES 52 – 55. 52 – 53 View FIGURES 56 – 63. 56 – 59 View FIGURES 64 – 68 )
Prionina simplex Casey, 1912: 252 View in CoL ; Knaus, 1914: 91; Lingafelter et al., 2014: 320 (type).
Prionus (Homaesthesis) simplex View in CoL ; Linsley, 1962: 53; Hovore & Turnbow, 1984: 4; Chemsak et al., 1992: 22 (checklist); Monn & Giesbert, 1994: 16 (checklist); Monn, 1995: 57 (cat.); Chemsak, 1996: 123; Heffern, 1998b: 174 (distr.); Monn & Hovore, 2005: 21 (checklist); 2006: 20 (checklist); Özdikmen & Turgut, 2009: 410; Bezark & Monné, 2013: 27 (checklist).
Prionus palparis View in CoL ; Lameere, 1919: 139 (syn.).
Prionus (Homaesthesis) rhodocerus Linsley, 1957: 10 View in CoL ; Linsley, 1962: 53; Hovore, 1981: 457 (distr.); Hovore & Turnbow, 1984: 4 (key); Chemsak et al., 1992: 22 (checklist); Monné & Giesbert, 1994: 16 (checklist); Monné, 1995: 57 (cat.); Chemsak, 1996: 122; Heffern, 1998: 6 (distr.); Monné & Hovore, 2005: 21 (checklist); 2006: 20 (checklist); Özdikmen & Turgut, 2009: 410; Bezark & Monné, 2013: 27 (checklist). Syn. nov.
Prionus (Homaesthesis) linsleyi Hovore, 1981: 455 View in CoL ; Hovore & Turnbow, 1984: 4 (key); Chemsak et al., 1992: 22 (checklist); Monné & Giesbert, 1994: 16 (checklist); Monné, 1995: 57 (cat.); Chemsak, 1996: 124; Monné & Hovore, 2005: 21 (checklist); 2006: 19 (checklist); Özdikmen & Turgut, 2009: 410; Bezark & Monné, 2013: 27 (checklist). Syn. nov.
Male ( Figs. 54-68 View FIGURES 52 – 55. 52 – 53 View FIGURES 56 – 63. 56 – 59 View FIGURES 64 – 68 ). Integument from reddish-brown (usually with parts brown or dark-brown) to dark-brown; antennae from reddish to dark-brown, always with basal segments darker; tarsi from reddish (usually with metatarsomere I darker) to dark-brown.
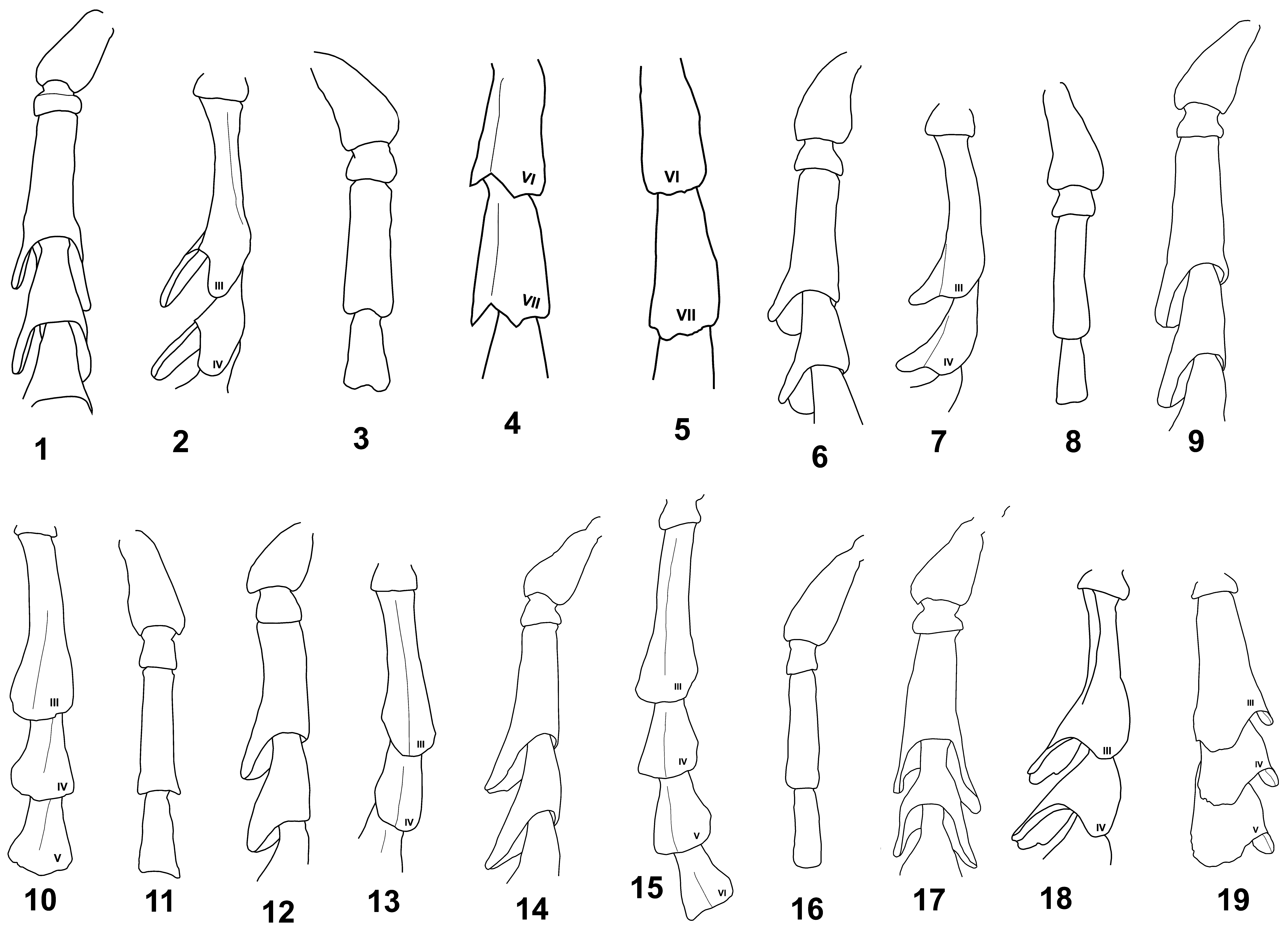
Head, excluding mandibles, from 0.8 to 1.1 times as long as prothorax at central area, elongate behind eyes (distance from posterior ocular edge to the prothorax from equal to it to longer than greatest length of upper eye lobe). Longitudinal dorsal furrow distinct from clypeus to near prothoracic edge (occasionally reaching anterior margin of prothorax), always more distinct from clypeus to posterior ocular edge. Area on each side of longitudinal sulcus coarsely, deeply, confluently punctate, usually more anastomosed near upper eye lobes and sparsely punctate on central area between posterior eyes and prothorax; dorsal surface with short, very sparse setae (sometimes slightly long and more conspicuous); area behind eyes moderately coarsely, punctate (usually somewhat vermiculate behind lower eye lobe), with long, moderately sparse setae behind upper eye lobes, more abundant behind lower eye lobes. Distance between antennal tubercles variable: from nearly contiguous to distinctly apart; coarsely, abundantly punctate on base, gradually sparser, finer toward apex, which usually is smooth. Postclypeus centrally narrowed, with or without transverse, deep sulcus near frons; posterior one-third distinctly, abruptly oblique toward anteclypeus (sometimes uniformly oblique); laterally coarsely, abundantly punctate laterally, with long, moderately abundant setae (often short and sparse); from smooth to moderately coarsely, abundantly punctate centrally, usually glabrous. Anteclypeus with anterior edge distinctly concave (sometimes slightly concave); shining, glabrous and impunctate. Labrum almost flat; with long, abundant setae laterally and near anterior edge. Eyes proportionally large, but upper eye lobes moderately narrow; distance between upper eye lobes from 1.0 to 1.3 times length of scape; distance between lower eye lobes from 1.2 to 1.7 times length of scape. Submentum trapezoid, slightly depressed; coarsely, confluently, shallowly punctate; with moderately short, sparse setae (distinctly sparser centrally); anterior edge distinctly carinate. Apex of labial palpi nearly attaining middle of maxillary palpomere IV (sometimes slightly surpassing). Mandibles (closed) from slightly shorter to slightly longer than 0.5 times length of head; latero-basal one-third with depression at inferior one-half (sometimes the depression almost reaches distal one-third); outer distal one-third strongly curved, forming an obtuse angle with basal two-thirds. Antennae with 12 segments; slightly surpassing middle of elytra. Scape, at most, attaining posterior ocular edge, enlarged toward apex; dorsally and laterally finely, sparsely punctate (laterally sparser); ventral surface very finely and sparsely punctate (sometimes almost smooth). Antennomere III (including distal projection) from 1.6 to 2.0 times longer than scape, distinctly enlarged toward apex (distal width at apex of projection from 1.6 to 2.0 times basal width); on dorsal view, imbrication distinct and projected ( Fig. 12 View FIGURES 1 – 19 ); apex of imbrication not emarginated ( Fig. 13 View FIGURES 1 – 19 ). Antennomere IV from 0.6 to 0.8 times as long as III; dorsally microsculptured and finely, sparsely punctate (sometimes only finely punctate toward inner side); imbrications as in III. Antennomeres V–XI dorsally microsculptured; imbrications as in III. Antennomere XII simple, without projections, flattened laterally. Antennomeres III–XII moderately carinate ventrally; imbrications from moderately to distinctly thickened; apex of imbrications from slightly to distinctly enlarged (mainly at antennomere XI).
Maximum prothoracic width from 0.7 to 0.8 times width of elytral base; anterolateral angle from distinct to absent ( Figs. 66–67 View FIGURES 64 – 68 ); middle lateral tooth acute, very distinct; posterolateral angle almost in a right angle, sometimes subrounded; margin between lateral middle tooth and posterolateral angle from slightly convergent to parallel; basal margin sinuous; anterior margin from concave to almost straight. Pronotum centrally subflat, frequently with slightly depression; not strongly explanate laterally; callosities distinct, but not strong; disc moderately abundantly punctate, but size and concentration of punctures highly variable throughout; with short, very sparse setae (sometimes absent). Prosternum finely asperate; with long, abundant setae (sometimes shorter throughout or toward prosternal process. Prosternal process not longitudinally sulcate; with short, abundant setae, longer at apex. Elytra moderately coarsely, abundantly punctate (general appearance can be somewhat rugose); each elytron with three carinae, fused at distal one-fourth (sometimes only two are fused); sutural spine short (sutural angle can be only projected). Metasternum microsculptured, less so toward metasternal suture; with long, abundant setae throughout. Metepisterna with sculpture and setae as those of metasternum laterally.
Ventrites I–IV microsculptured, finely, sparsely punctate; ventrite I–IV with short, moderately abundant setae (sometimes somewhat longer on ventrite I). Tarsomeres I–III moderately slender (mainly metatarsomeres); pro- and mesotarsomeres I–III acute at apex (sometimes only tarsomeres I–II); metatarsomeres I–III spined at apex; pro- and mesotarsomeres I–III with spongy setal pads on ventral surface, with narrow subglabrous longitudinal area centrally (sometimes slightly wider); metatarsomeres I–III with spongy setal pads distinctly less conspicuous and with central longitudinal sulcus distinctly wider than on pro- and mesotarsomeres; metatarsomere I about as long as II–III together.
Female. Not examined.
Dimensions in mm (male). Total length (including mandibles), 18.6–26.4; prothoracic length at center, 3.0– 4.3; greatest prothoracic width, 5.9–8.8; humeral width, 7.8–10.4; elytral length, 13.6–18.1. Dimensions according to Casey (1912): Length (♂) 22.5 mm.; width 9.0 mm.” According to Linsley (1957): “Length, 19–25 mm. [male]”; “Length, 22.5 mm. [female].” According to Hovore (1981): “ 15.5 mm [male]”; “ 20 mm [female].”
Geographical distribution. Prionina simplex was known only from U.S.A. (Kansas). Monné (1995) recorded: “Eastern Kansas and eastern Oklahoma.” According to Heffern (1998b): “ Alexander (1958) reported P. simplex from two locations in Craig County, Oklahoma. Alexander’s paper appears to be the basis for two localities shown in northeastern Oklahoma on the distribution maps given by Linsley (1962) and Chemsak (1996). The one locality shown in central Kansas on the map by Chemsak (1996) represents just a “state record” and not a specific collecting record (J. Beierl, Editor of Wolfsgarden Books, pers. comm.)”; “ In 1997, the author borrowed three specimens identified as P. simplex from the K. C. Emerson Museum at Oklahoma State University, Stillwater, OK through the courtesy of its curator, Dr. D. C. Arnold. These specimens, all from Craig Co., bear determination labels from J. N. Knull in 1937. However, all of these specimens are P. (Neopolyarthron) debilis Casey ”; and “The type specimen appears to be the only known example of this species.” However, Alexander (1958) did not report two localities in Craig County, he only pointed out: “ Prionus simplex Casey. Craig (OSU).” Nevertheless, following Heffern (1998b), the inclusion of P. simplex in Oklahoma by Linsley (1962) and Chemsak (1996) was a mistake.
The geographical distribution including those of P. rhodocerus and P. linsleyi are: USA [Utah ( Linsley, 1957), Kansas ( Casey, 1912), Nebraska ( Linsley, 1957); Colorado ( Hovore, 1981), New Mexico ( Hovore, 1981); Arizona ( Hovore, 1981); Wyoming (Chemsak, 1996)].
Type, type localities. Of Prionina simplex ( Figs. 54–55 View FIGURES 52 – 55. 52 – 53 ): Holotype male, from Kansas, deposited at USNM. Figured at Lingafelter et al. (2016).
Of Prionus (Homaesthesis) rhodocerus ( Figs. 60–63 View FIGURES 56 – 63. 56 – 59 ): Holotype male from USA (Utah, Red Creek) deposited at CAS. Three paratypes from Utah, deposited at CAS and USNM. Figured at Bezark (2016).
Of Prionus (Homaesthesis) linsleyi ( Figs. 56–59 View FIGURES 56 – 63. 56 – 59 ): Holotype male and paratype female from USA (Arizona, “Coconimo County, 6.5 mi S. Moenkopi”), deposited at CAS. Figured at Bezark (2016).
Material examined. USA, no other data, 1 male ( USNM). Arizona: Chino Valley, 1 male, 19.VII.1939, [no collector indicated] ( ESSIG). New Mexico: Quay County (sand dunes, 1.5 mi. N San Jon), 1 male, V.20.1986, P. A. Opler col. ( ESSIG). Utah: Fernow Valley, Juab County, 1 male, 03.VIII.1939, [no collector indicated] ( USNM); Fort Duchesne, 1 male, 07.XII.1932, Lowell Cutler col. ( USNM); Little Granite Mountain (Dugway Proving Grounds, Tooele County), 1 male, 24.VIII.1953, M. D. Baxter col. ( ESSIG).
Remarks. Hovore (1981) recorded: “ Prionus (H.) linsleyi is closely related to P. (H.) simplex (Casey) , from the midwest, and P. (H.) rhodocerus Linsley , from the Rocky Mountain states. Based upon a comparison of holotype males, linsleyi differs from simplex by the shape of the scutellum (sides evenly rounded to apex in simplex ), more pronounced elytral sculpture, longer, denser metasternal pubescence (finer, shorter and sparser in simplex ), and by the longer, parallel-sided first posterior tarsal segment (3 times as long as the apical width in linsleyi , subtriangular in dorsal outline and less than twice as long as apical width in simplex ).” Based on species with a large amount of specimens examined, we can affirm that the shape of the scutellum is notably variable in Prionus . The elytral sculpture is somewhat different in P. simplex (finer) and P. linsleyi (coarser). But, as it is possible to see in other species, the elytral sculpture is frequently variable. The metasternal pubescence could be a good difference among those species. However, metasternal pubescence is highly variable in other species of the genus and, in this case, it is not too different in both holotypes. Regarding the first metatarsomere: it is about twice as long as the apical width in P. s i m pl ex. Studying photographs of the holotype of P. linsleyi indicates the first metatarsomere is slightly longer than twice its apical width. Note: we have found that the width of first metatarsomere is frequently variable in species of Prionus .
Hovore & Turnbow (1984) separated P. sim pl ex and P. linsleyi from P. spinipennis and P. rhodocerus in the couplet 5: “Coloration dark brown, castaneous, or piceous; antennae all or in part lighter reddish-brown”— P. spinipennis and P. rhodocerus ; “Coloration concolorous light reddish-brown, antennae not contrasting”— P. simplex and P. linsleyi . However, the holotype of P. simplex , and the specimens examined are not light reddishbrown [they are dark-brown—“evenly red-brown, according to Casey (1912)], and the antennae, at least in one specimen, are in part distinctly contrasting [“the antennae paler and more ochraceous”, according to Casey (1912)]. Linsley (1962) and Chemsak (1996) recorded on the color: “Integument red-brown throughout.” The color in specimens of Prionus is frequently variable, including legs and antennae, as we could see examining specimens of P. emarginatus . Thus, the color is not useful in separating species of Prionus .
Lameere (1919), in doubt, synonymized, P. simplex under P. pa l pa r i s. Linsley (1962), and all other authors who published on P. simplex , apparently, did not observe this doubtful synonym. We agree with Linsley (1962): P. simplex is not equal to P. palparis . The distance between upper eye lobes in the latter is distinctly smaller.
We found an interesting variation in the P. simplex specimens examined by us as the lateroanterior angle of pronotum may be absent (as in the holotype of P. linsleyi ) or distinct (as in holotype of P. simplex and P. rhodocerus ). This variation invalidates the difference pointed out in the three keys that included P. simplex . According to Linsley (1962): “anterior lateral angle of pronotum distinct or dentiform”—to P. palparis , P. simplex , and P. rhodocerus ; “anterior lateral angle of pronotum rounded or indistinct”—to P. integer . In Chemsak (1996): “Pronotum with anterior angle distinct”—to P. simplex ; “Pronotum with anterior lateral angle irregularly rounded”—to P. linsleyi . And according to Hovore & Turnbow (1984): “Anterior pronotal angle distinct”—to P. simplex ; “Anterior lateral pronotal angle irregularly rounded”—to P. linsleyi .
According to Linsley (1957): “The specimens from Kansas and Nebraska have the integument, other than antennae, black rather than piceous, the metasternum thinly pubescent, the abdomen nearly glabrous, and the punctation of the pronotum and elytra finer. I have hesitated to designate them as subspecifically different in the absence of material from the intervening area.” We saw the same kind of variation in the specimens examined. Also, we noted that the projection of the antennomeres is highly variable in the specimens: it could be distinctly enlarged (including antennomeres X–XI), or distinctly slender (mainly in antennomeres X–XI). The antennomeres can also be distinctly tumid or less so.
Based on comparison of the specimens examined by us, original descriptions, and photographs of the holotypes with good resolution of details, we conclude that Prionus linsleyi and P. rhodocerus are synonyms of P. simplex , and the latter differs from P. integer only by the number of antennomeres (respectively: 12 and 13).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Prionus (Homaesthesis) simplex ( Casey, 1912 )
| Santos-Silva, Antonio, Nearns, Eugenio H. & Swift, Ian P. 2016 |
Prionus (Homaesthesis) linsleyi
| Bezark 2013: 27 |
| Ozdikmen 2009: 410 |
| Monne 2005: 21 |
| Monne 1994: 16 |
| Chemsak 1992: 22 |
| Hovore 1984: 4 |
| Hovore 1981: 455 |
Prionus (Homaesthesis) simplex
| Bezark 2013: 27 |
| Ozdikmen 2009: 410 |
| Monn 2005: 21 |
| Heffern 1998: 174 |
| Monn 1995: 57 |
| Monn 1994: 16 |
| Chemsak 1992: 22 |
| Hovore 1984: 4 |
| Linsley 1962: 53 |
Prionus (Homaesthesis) rhodocerus
| Bezark 2013: 27 |
| Ozdikmen 2009: 410 |
| Monne 2005: 21 |
| Monne 1994: 16 |
| Chemsak 1992: 22 |
| Hovore 1984: 4 |
| Hovore 1981: 457 |
| Linsley 1962: 53 |
| Linsley 1957: 10 |
Prionus palparis
| Lameere 1919: 139 |
Prionina simplex
| Lingafelter 2014: 320 |
| Knaus 1914: 91 |
| Casey 1912: 252 |