
Pinguicula sehuensis Bacch., Cannas & Peruzzi, 2014
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.186.5.5 |
|
persistent identifier |
https://treatment.plazi.org/id/8F3087AC-FFAA-F139-FF22-FBB1FF4CFD31 |
|
treatment provided by |
Felipe |
|
scientific name |
Pinguicula sehuensis Bacch., Cannas & Peruzzi |
| status |
sp. nov. |
Pinguicula sehuensis Bacch., Cannas & Peruzzi View in CoL sp. nov. ( Fig. 1 View FIGURE 1 ).
Type:— ITALY, Sardinia. Montarbu, Seui , calcari mesozoici, 1160 m a.s.l., 39° 53’ N – 9° 23’ E, 3 May 2014, G GoogleMaps . Bacchetta & M . Marras s.n. (holotype: CAG!; isotypes: PI! FI!) .
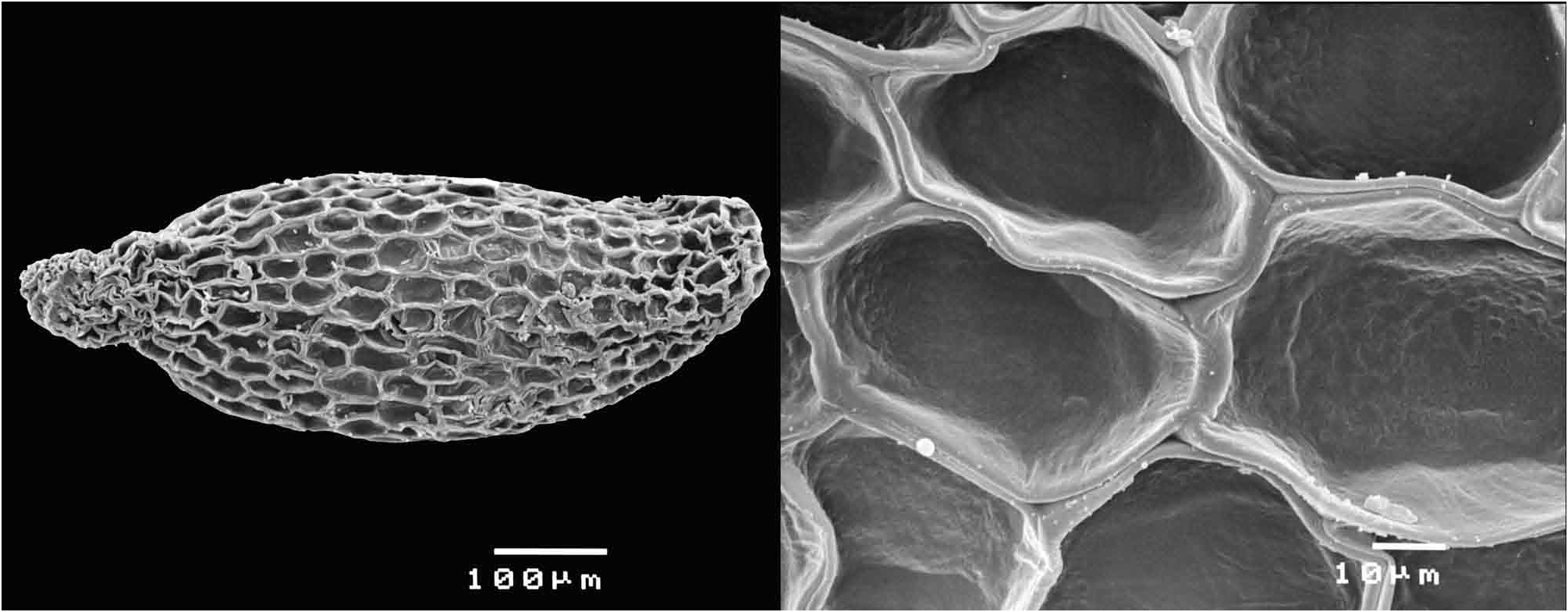
Perennial herb, small, rosette forming, scapose, succulent. Stem short, with ascending, not branching rhizome and numerous adventitious fibrous roots. Rosettes with few, 4 – 5(– 6) leaves lying more or less flat on the ground, homophyllous. Overwintering as buds (hibernacula). Leaves in outline obovate-oblong-triangular, (15 –)20 – 30(– 35) mm long, (7 –)9 – 12(– 16) mm broad at anthesis. The margin entire, slightly incurved; brittle; the upper surface densely covered with mucilaginous sessile and stalked glands. Scapes 1 – 2(– 4), erect, (16 –)30–50(– 70) mm tall, terete, 1- flowered, directly beneath the flower densely covered with stipitate glands, to the base sparsely glandular. Flowers (18.5 –)19 – 24.5(– 24.9) mm long (spur included). Spur slightly curved downwards, thin, (6.1 –)6.5 – 9.5(– 9.7) mm long. Corolla distinctly bilabiate, lilac violet, white at the centre, at the throat white with violet veins, spur violet; throat densely covered with white clavate hairs; upper lip with 2 nearly identical lobes not overlapping, (5.9 –)6.2 – 8(– 8.2) mm long and (4 –)4.2 – 5.2(– 5.4) mm broad, erect to patent, apex rounded; lower lip much larger than the upper lip, with 3 lobes not overlapping, lateral lobes (6.9–)7.4–9.2(–9.6) mm long and (4.8–)5.4–6.1(–6.3) mm broad, median lobe (7.4–)7.7–9.3(–9.5) long and (6.1–)6.3–7.5(–7.8) mm broad, apex rounded, the whole corolla appears well open [opening angle (110 –)120 – 160°]. Calyx distinctly bilabiate, densely covered on both surfaces and the margins with stipitate glands, (3.3 –)3.5 – 4(– 4.2) mm in total diameter, upper lip divided nearly to 1/2 – 2/3 into 3 lobes elliptic or more rarely elliptic-triangular; lower lip divided to 1/2 into 2 short lobes. Capsule subglobose-ovoid, (2.7 –)2.8 – 3.3(– 3.4) mm long, (2.2)2.5 – 3(– 3.1) mm in diameter, 1-loculate. Seeds minute, like sawdust, with reticulate surface, elliptic, 500 – 700 µm long, 200 – 300 µm wide ( Fig. 2 View FIGURE 2 ). Ratio micropylar seed appendage / seed length: 0.11 – 0.16. Number of exotesta cells 250 – 450, exotesta cells 25 – 70 µm long, 12.5 – 50 µm wide; depth of anticlinal walls of exotesta cells 3 – 8.5 µm. Cotyledon 1, convolute, asymmetric. Pollen spheroidal, 4 – 6(– 7) colporate, (25 –)30 – 35(– 40) µm. Flowering ( IV) V – VI; fruiting VI – VII.
Etymology:—The specific epithet refers to “Sehui“, the ancient name of the town and white mount of Seui (Ogliastra, Sardinia).
Ecology:— Pinguicula sehuensis is a chasmophyte linked to calcareous substrates as limestone, travertine and conglomerate, where it grows preferentially on vertical cliffs at an altitude between 1035 and 1185 m a.s.l. It is especially linked to subalkaline, scarcely developed, soils. It grows on rocks preferentially exposed to NW-N (312–326°) under a pluviseasonal Mediterranean bioclimate, within the upper meso-Mediterranean and lower supra-Mediterranean belts, with a upper subhumid and lower humid ombrotype ( Bacchetta et al. 2009). It is a member of chasmohygrophilous plant communities characterized by Arenaria balearica L., Bellium bellidioides L., Potentilla caulescens L. subsp. nebrodensis (Strobl ex Zimmeter) Arrigoni , Saxifraga pedemontana All. subsp. cervicornis (Viv.) Engl. and Sesleria barbaricina (Arrigoni) Arrigoni. Other hygrophytes like Platanthera algeriensis Batt. & Trab. , Samolus valerandi L., Schoenus nigricans L. and Solenopsis bivonae (Tineo) M.B.Crespo, Serra & Juan also occur in the same habitat.
Karyology:—The population from the locus classicus resulted with 2 n = 16 chromosomes ( Fig. 3 View FIGURE 3 ). The chromosomes are very small, ranging from 0.8 to 1.2 μm in length.
Distribution:—On the basis of our field investigations, this species appears quite rare and scattered just along the North-Western part of Montarbu ( Fig. 4). Currently, only eight metapopulations are known, seven of them occur in vertical cliffs (chasmophytic habitat) and one is linked to small terraces among cliffs (comophytic habitat).
Conservation:—Six of the eight populations are threatened by goat and mouflon grazing, albeit a metapopulation decline was not observed. Considering the small population size and the potential increase of threat level (due to human activity or stochastic events), we assess here P. sehuensis as Vulnerable (VU D2), by applying the IUCN red listing protocols ( IUCN 2001). The total number of mature plants ranges from 250 to 1000, distributed in less than 10 km 2. The Centro Conservazione Biodiversità (University of Cagliari) started a conservation programme at the Sardinian Germplasm Bank (BG-SAR), including a monthly monitoring of all metapopulations.
Taxonomic relationships:—Other species of P. sect. Pinguicula showing some resemblance with P. sehuensis are P. dertosensis ( Cañigueral 1957: 413) Mateo & Crespo (1995: 430) , P. grandiflora Lam. and P. corsica Bernard & Gren. in Grenier & Godron (1853: 443). P. dertosensis shows very distinctive ovoid seeds ( Zamora et al. 1996, under the name P. submediterranea Blanca, Jamilena, Ruíz Rejón & Reg.Zamora in Zamora et al. 1996: 57), larger and more numerous leaves, and longer scapes, while P. grandiflora has larger, more intensely coloured flowers ( Blanca et al. 1999; Blanca 2001). Pinguicula corsica has mainly whitish-blue flowers with overlapping lobes, a short spur, (3 –)4 – 6(– 9) mm long ( Casper 1966), a plicate-symmetric cotyledon ( Degtjareva et al., 2004) and a different ecology (subalpine/alpine belts, see Contandriopoulos 1962). The seed features of P. sehuensis are in general accordance with those typical of other species within P. sect. Pinguicula . In particular, the convolute aestivation of the cotyledon was so far evidenced only in P. grandiflora ( Degtjareva et al. 2004) . However, no detailed data are available concerning the seeds of P. dertosensis and other taxa endemic to the Iberian Peninsula such as P. nevadensis ( Lindberg 1932: 142) Casper (1962: 112) . It is interesting to note that P. dertosensis (the species morphologically most similar to P. sehuensis ) is generally regarded as systematically close to P. grandiflora ( Blanca et al. 1999, Blanca 2001). Concerning karyology, 2 n = 16 is commonly regarded as the diploid status in P. sect. Pinguicula (Peruzzi 2004) , and this number was so far reported only for other two European species: P. corsica ( Contandriopoulos 1962) and P. nevadensis ( Zamora et al. 1996) . Zamora et al. (1996) hypothesized that P. dertosensis , which they regarded as a hexaploid with 2 n = 48 chromosomes, had an allopolyploid origin, involving P. grandiflora (2 n = 32) and an unknown diploid species. In this framework, P. sehuensis could indeed represent this diploid species. However, it must be noted that Casper & Stimper (2009), in their survey on Pinguicula chromosome numbers, while confirming the diploid status of P. corsica , actually confuted the counts for P. dertosensis and P. nevadensis , which they reported as octoploid with 2 n = 64 and tetraploid with 2 n = 32 chromosomes, respectively. Irrespective of this, we can consider P. sehuensis as a patroendemic species, i.e. a diploid unit which has only polyploid relatives, similarly to P. corsica ( Siljak-Yakovlev & Peruzzi 2012) . However, P. corsica and P. sehuensis , albeit both diploid and geographically close, seem not obviously related each other, neither on taxonomical nor on ecological grounds. A molecular phylogenetic study of all central Mediterranean endemic taxa is planned and will clarify the evolutionary relationships of P. sehuensis with other species of P. sect. Pinguicula .
Additional specimens seen (paratypes):— ITALY. Sardinia: Montarbu, Seui ( Ogliastra ), calcari mesozoici, 1035 m, NW 315°, 39°53’N – 9°21’E, 24 June 2012, G GoogleMaps . Bacchetta s.n. ( CAG!); Pirastu Trottu, Montarbu , Seui ( Ogliastra ), rocce stillicidiose su travertino, 1185 m, N 15 °, 39°53’N – 9°23’E, 19 June 2013, G GoogleMaps . Bacchetta & A . Delage s.n. ( CAG!) .


| G |
Conservatoire et Jardin botaniques de la Ville de Genève |
| M |
Botanische Staatssammlung München |
| CAG |
Università degli Studi di Cagliari |
| PI |
Paleontological Institute |
| FI |
Natural History Museum |
| V |
Royal British Columbia Museum - Herbarium |
| VI |
Mykotektet, National Veterinary Institute |
| A |
Harvard University - Arnold Arboretum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |