
Patuloricus tangaroa gen
|
publication ID |
https://doi.org/ 10.1016/j.jcz.2022.06.004 |
|
publication LSID |
lsid:zoobank.org:pub:D67E1882-834D-469A-B034-B31D4C799533 |
|
persistent identifier |
https://treatment.plazi.org/id/03D887CB-FFE3-C507-FFF9-FDF7FA60FA54 |
|
treatment provided by |
Felipe |
|
scientific name |
Patuloricus tangaroa gen |
| status |
|
3.2. Patuloricus tangaroa gen et sp. nov
( Figs. 6–9 View Fig View Fig View Fig View Fig ; Table 1). urn:lsid:zoobank.org:act:C238E126-CC58-4489-91D3-
E1EEE2B2F42D urn:lsid:zoobank.org:act:8B29532C-6212-44FE-84EA-
4E8D42B2680B Taxonomic account. Phylum Loricifera Kristensen, 1983. Family incertae sedis.
3.2.1. Patuloricus gen. nov. diagnosis
Higgins larvae belonging to the Shira larva type. Shira larva with large, bulbous head. Mouth cone without inner armature; introvert with six scalid rows. Trunk plicated, consisting of long, accordion-like thorax, and short abdomen. Thorax region about twice as long as abdomen. Thorax with anterolateral and anteroventral setae; abdomen with posterodorsal and posterolateral setae. Toes long and slender, but not acicular. Postlarva with large, globular head. Mouth cone with inner armature. Head with a few longitudinal retractor muscles. Trunk, slender and elongate, densely packed with numerous longitudinal and circular muscles.
3.2.2. Etymology
The new genus is named Patuloricus . The prefix, Patu-, is derived from the Maori ¯weapon of the same name, which is club-shaped and very similar to the shape of the Shira larva. The name is furthermore a dedication to the indigenous M¯aori population of New Zealand. The postfix, - loricus, appears in all Loricifera genus names, and refers to the presence of a lorica.
3.2.3. Patuloricus tangaroa sp. nov. diagnosis
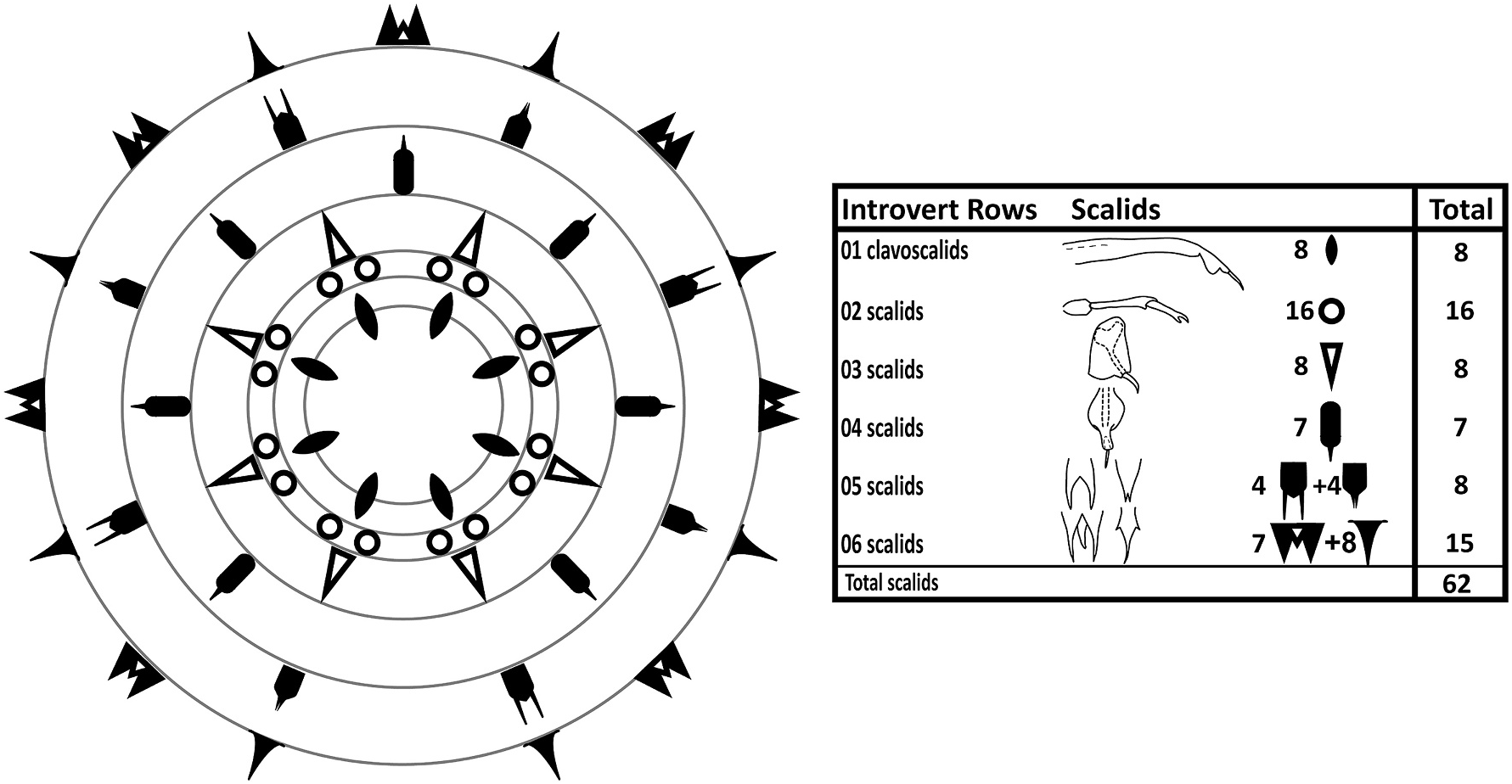
Species with diagnostic traits of Patuloricus gen. nov. Shira larva with introvert equipped with six scalid rows. Introvert Row 1: eight clavoscalids, composed of long proximal unit, lobed medial unit, and acicular end-piece. Introvert Row 2: sixteen scalids composed of short basis, elongate medial unit, and end-piece with pincer-shaped claw. Introvert Row 3: eight scalids composed irregular rectangular plate, and a hook-like tip. Introvert Row 4: seven scalids composed of a plate-like unit extending into a shaft, holding a distal, acicular tip. Introvert Row 5: four bifurcating scalids alternating with four scalids forming triangular plates with bifurcated tips. Introvert Row 6: seven trifurcating scalids alternating with eight kite-shaped scalids. Accordion-like thorax with more than 30 elongate plicae, and eight transverse constrictions. Abdomen with 24–28 deeper plicae, but no transverse constrictions. Simple, acicular, curved to curled anterolateral and anteroventral setae present on thorax. Abdomen with long posterodorsal setae with bulbous bases, and much shorter and stout posterolateral setae. Toes slender, tapering abruptly about halfway from tips, and again at their pointed, offset tips.
3.2.4. Etymology
The new species is named tangaroa , after the Polynesian Tangaroa – the Atua (God or spirit) of oceans, lakes, and rivers, and all aquatic creatures.
3.2.5. Examined material
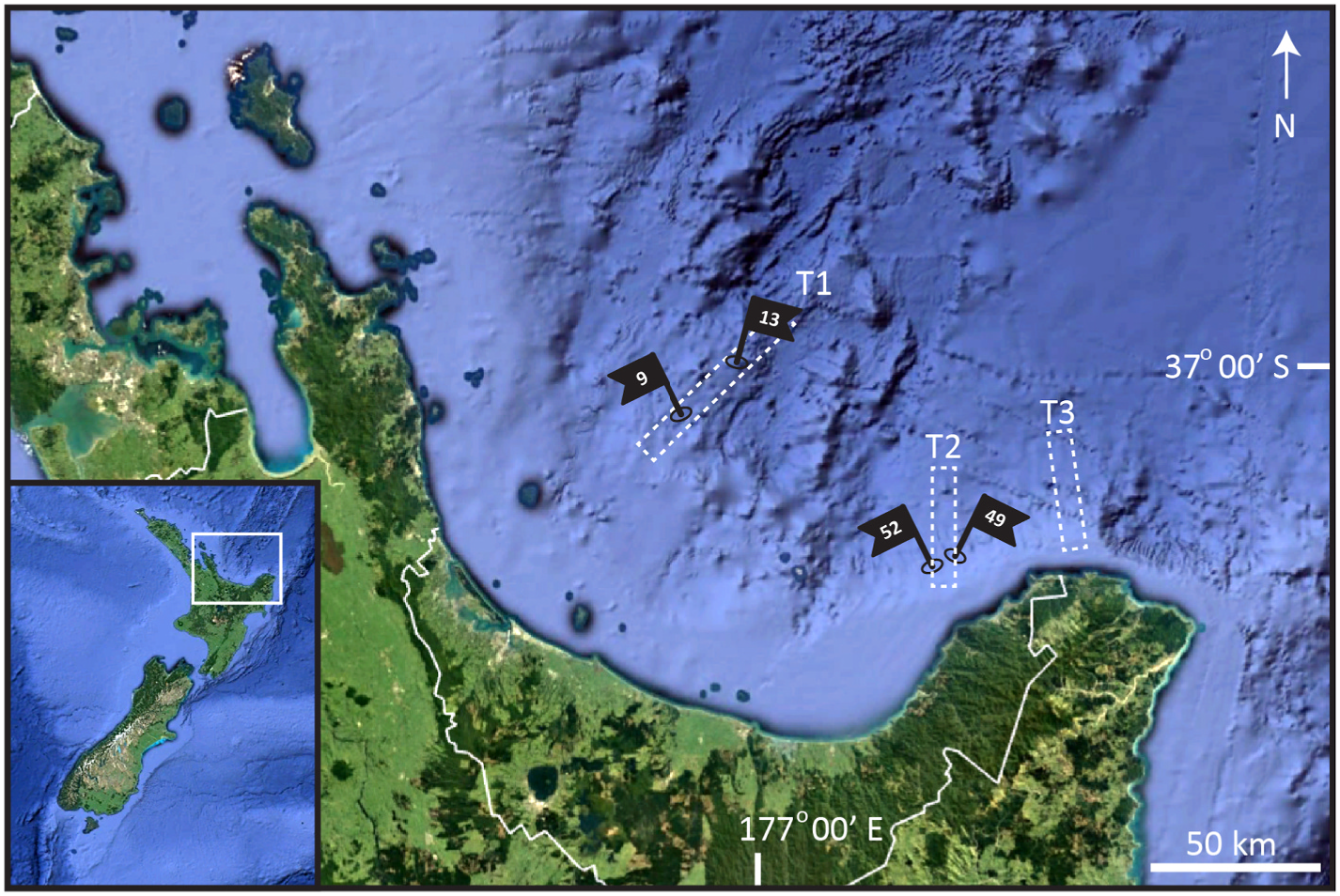
Holotype, Shira larva containing a postlarva, collected from silt in April 2012, at St. 52 (sensu Rosli et al. 2016) at 710 m depth on the continental slope, in the eastern part of Bay of Plenty, North Island, New Zealand (37 ◦ 30.26’N, 177 ◦ 37.19’E), mounted in glycerine, and deposited at NIWA under catalogue number NIWA-159433. See Fig. 1 View Fig for localities and Table 1 for detailed station data. GoogleMaps
3.2.6. Description of Shira larva
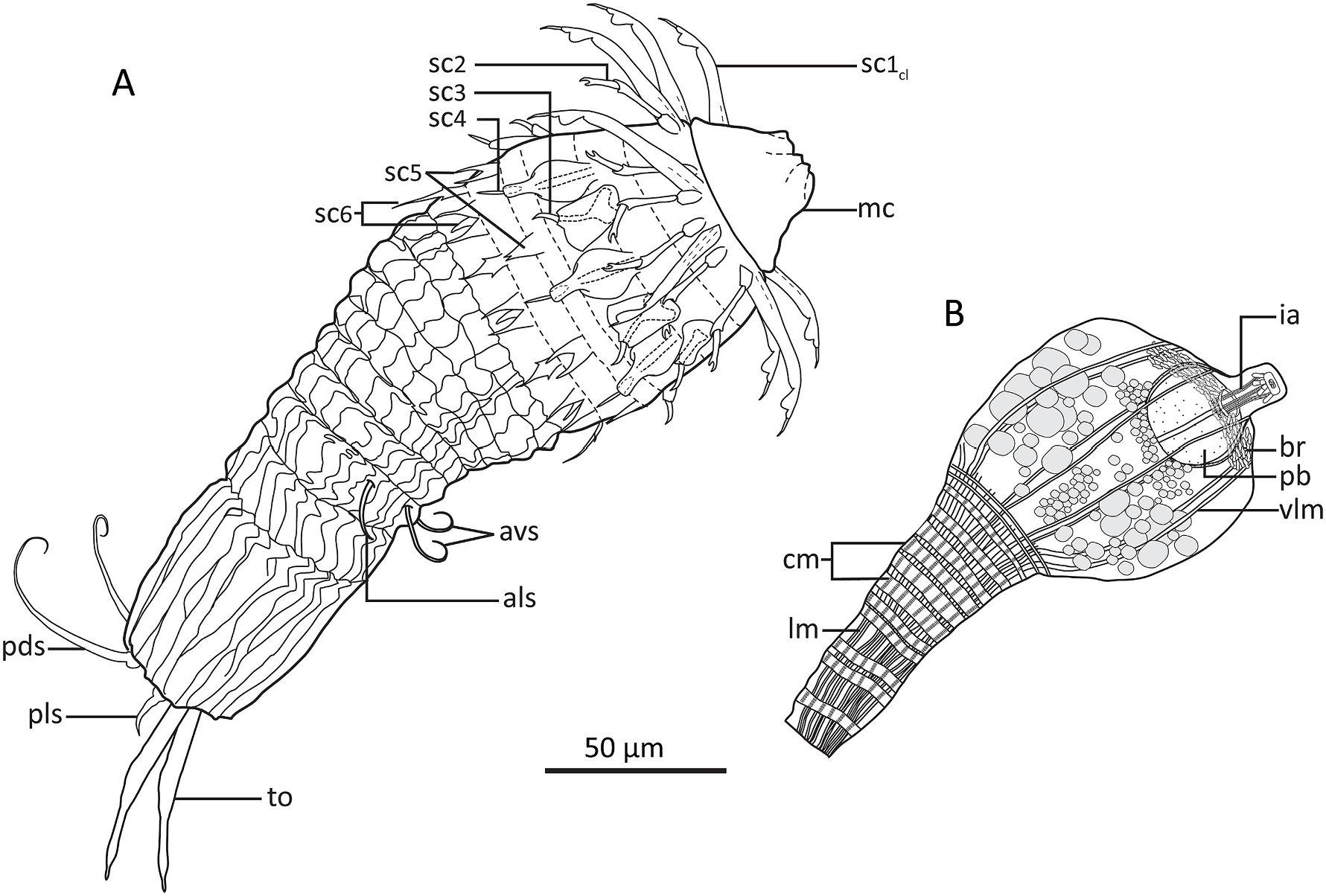
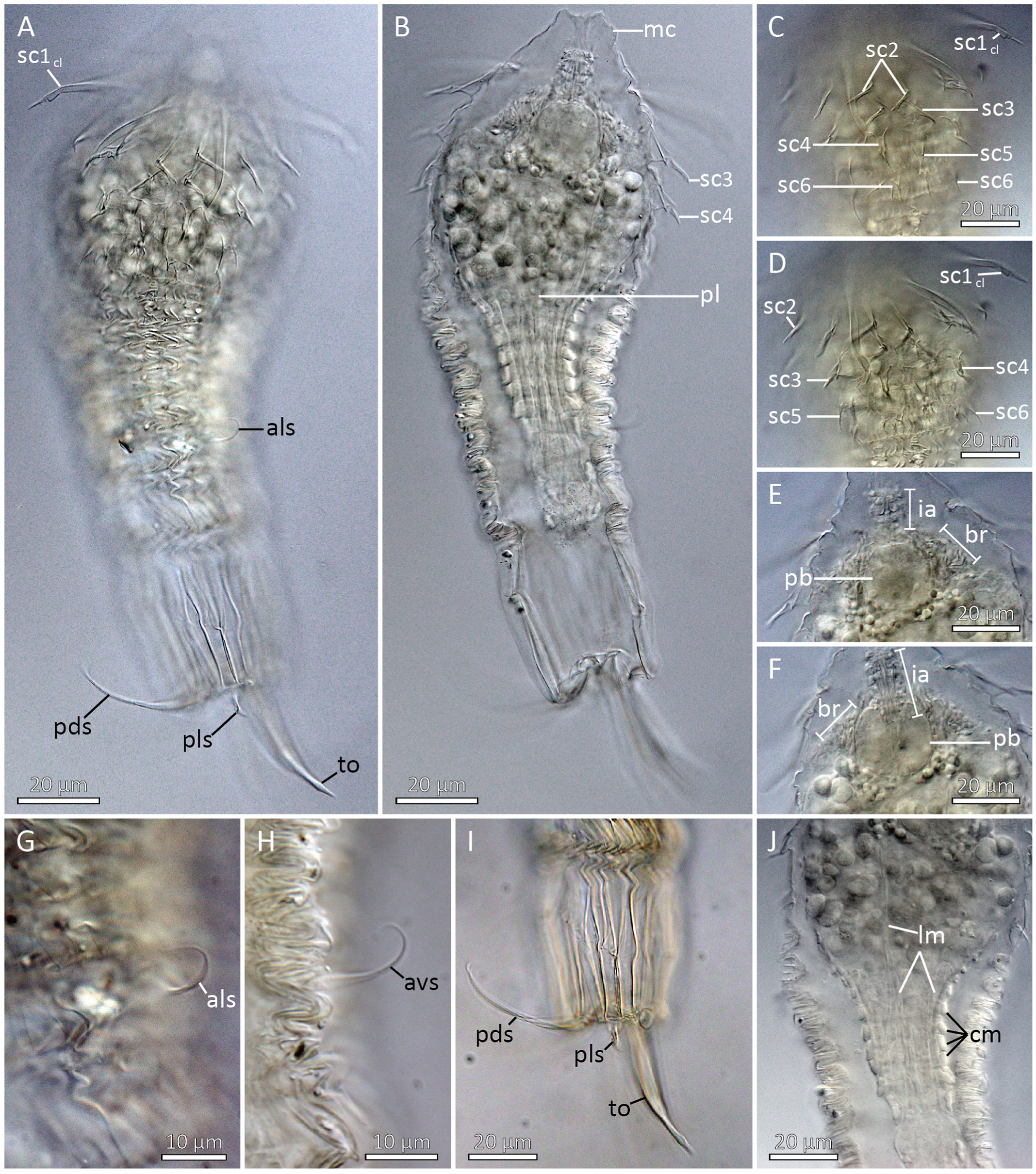
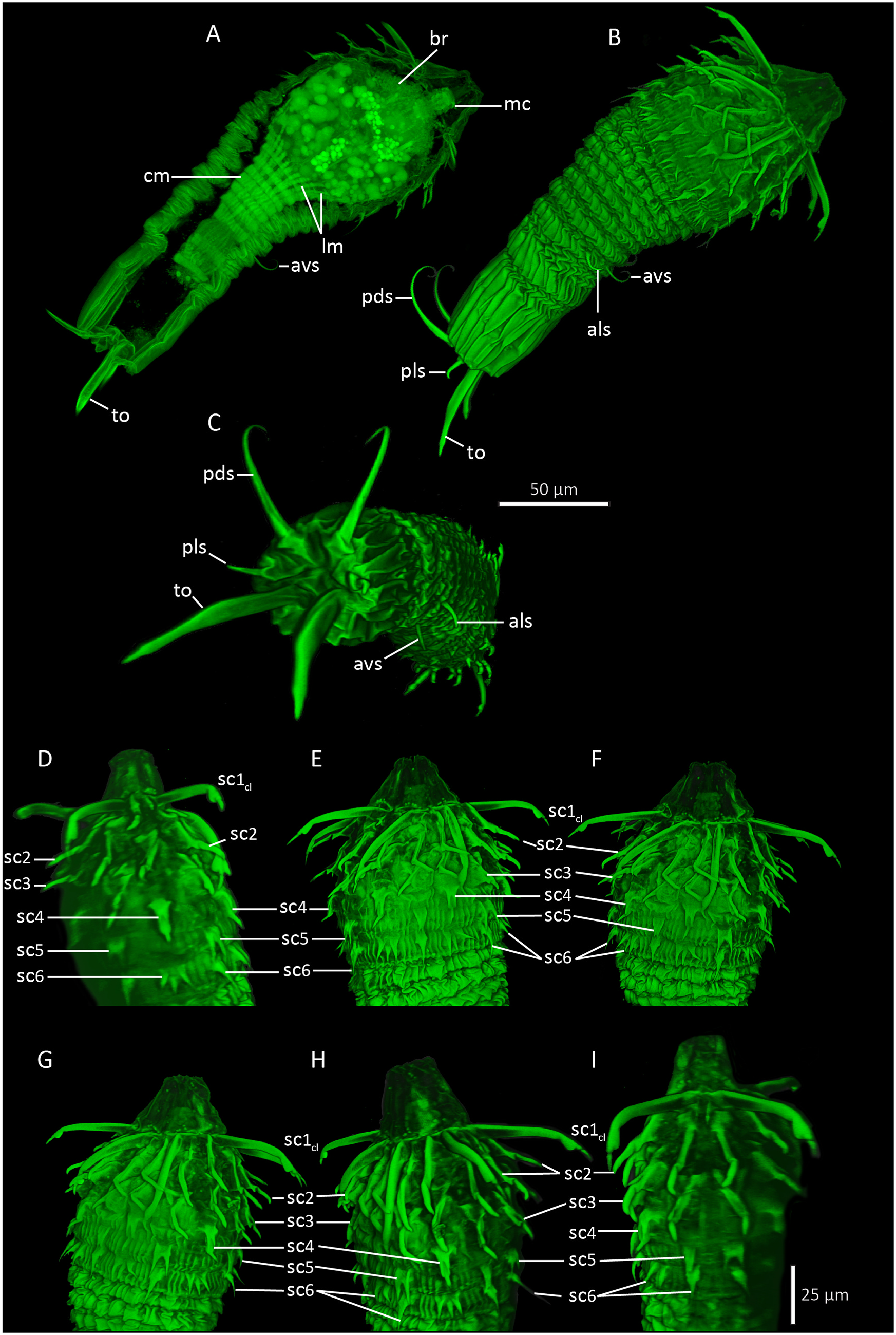
Holotypic Shira larva ( Figs. 6A View Fig , 7 View Fig and 8 View Fig , 9 View Fig ) measures 218 μm in length, inclusive mouth cone. The head is round and bulbous, measuring 75 μm at it widest point, whereas the trunk is more slender, measuring 63 μm at its widest point, i.e., near the anterior lorica margin, and only 40 μm more posteriorly. The mouth cone is broadly conical, with a wide base (40 μm) and a narrower tip (16 μm). No internal structures are present in the mouth cone ( Fig. 8B View Fig ).
The introvert has six rows of scalids ( Fig. 7 View Fig ). The anteriormost Row 1 consists of eight clavoscalids. The clavoscalids are about 47 μm long, and are composed of three units: a proximal smooth and flattened part representing more than 3/4 of the total scalid length, a short, lobed middle piece, and an even shorter acicular end-piece. All eight clavoscalids show a uniform morphology, and they attach bilaterally symmetrical with four on each side of the introvert, but none in middorsal or midventral positions ( Figs. 6A View Fig , 7 View Fig and 8A View Fig , C-D, 9B, D-F).
Row 2 consists of sixteen scalids that attach pairwise at the bases of the clavoscalids. Each scalid (length: 25 μm) consists of a short basis, an elongate mid-piece, and short end-pieces with an arm terminating into a small, pincher-shaped claw ( Figs. 6A View Fig , 7 View Fig , 8 View Fig C-D, 9B, D-F).
Row 3 consists of eight scalids that are longitudinally aligned with the clavoscalids. Each scalid consists of a large, irregular rectangular plate that attaches to the introvert, and a hook-like tip (length: 10 μm) that projects from each plate ( Figs. 6A View Fig , 7 View Fig , 8 View Fig B-D, 9B, D-F).
Row 4 consists of seven scalids, with one scalid attaching middorsally and the remaining six as three bilateral symmetrical pairs. Like those in Row 3, the scalids are composed of a plate-like element that attaches to the introvert, but this plate is more oval, and extends distally into a shaft. A short, distal, straight, acicular tip (length: 9 μm) emerges from the shaft, like a pen nib fitting into a pen holder ( Figs. 6A View Fig , 7 View Fig , 8 View Fig B-D, 9B, D-F).
Row 5 consists of four + four dimorphic scalids that appear alternatingly around the introvert. Four scalids (length: 14 μm) have a deep, median cleft, thus forming bifurcated structures, whereas another four are small triangular plates (length: 10 μm), where only the distal tips show a very small bifurcation. The scalids in this row are longitudinally aligned with the clavoscalids and the scalids of Row 3, but as a result of their alternation between two different kinds, the scalids in this row are also the only ones in the introvert that do not appear in a bilateral symmetrical pattern ( Figs. 6A View Fig , 7 View Fig , 8 View Fig C-D, 9B, D-F).
Row 6 consists of seven plus eight dimorphic scalids that appear alternatingly around the introvert. Seven scalids (length: 16 μm) resemble the deeply bifurcating ones in Row 5, but they have an additional medial extension (although shorter than the lateral ones), which makes them trifurcated. The remaining scalids (length: 20 μm) are kite-shaped, with short lateral tips, and a long, slender distal tip. The seven trifurcated scalids are longitudinally aligned with the seven Row 4 scalids (thus, one is located in middorsal position), whereas the eight kite-shaped scalids are aligned with Row 1, 3 and 5 scalids. This arrangement makes the two scalid types alternate, and appear bilateral symmetrical ( Figs. 6A View Fig , 7 View Fig , 8 View Fig C-D, 9B, D-F).
A distinct neck region is not present. The trunk is much more slender than the bulbous head, and divided into a thorax and an abdomen ( Figs. 6A View Fig , 8 View Fig A-B, 9A-B). The entire trunk is covered by numerous plicae. The thorax has more than 30 plicae, and is characterized by about eight transverse constrictions, which gives it an accordion-like appearance. The partly contracted thorax is about 80 μm in length, making it almost twice as long as the abdomen. The thorax has two pairs of setal appendages that attach between the second and third posteriormost transverse constriction. The anterolateral setae (length: 16 μm) are simple and slightly curved, whereas the anteroventral ones (length: 22 μm) are more curled ( Figs. 6A View Fig and 8A View Fig , G-H, 9B–C).
The short abdomen (length: 50 μm) has fewer (about 24–28) but deeper plicae than the thorax. Cuticular surface patterns are restricted to longitudinal plicae, and there is no trace of any transverse constrictions ( Figs. 6A View Fig , 8 View Fig A-B, I, 9B–C). Abdominal appendages include two pairs of setae, and the paired toes. The posterodorsal setae are rather long (length: 58 μm) and distally curled, whereas the posterolateral setae are short (length: 9 μm), rigid and stout. Both the posterodorsal and posterolateral setae attach to small, bulbous bases. The toes (length: 48 μm) are slender, and divided into three parts. The proximal and thickest parts taper abruptly about halfway on the toes, and become conspicuously thinner. Towards the tips, these thinner distal parts get slightly thicker, until they again taper abruptly, forming short pointed toe tips ( Figs. 6A View Fig and 8A, I View Fig , 9 View Fig A-C).
3.2.7. Description of postlarva
The postlarva (length: 158 μm) is conspicuously pear-shaped, with a globular head and a slender trunk ( Figs. 6B View Fig , 8B View Fig and 9A View Fig ). Its body cuticle is so thin that it is impossible to observe any outer cuticular structures, except for the actual outlining of the postlarva.
The head measures ca 67 μm in both length and width, and has a short, apical mouth cone (length: 16 μm). Apparently, the head is completely devoid of scalids or any other outer, cuticular structures. Internal structures include inner armature of the mouth cone, a globular pharyngeal bulb, a ring-shaped brain, and at least four pairs of longitudinal muscles ( Figs. 6B View Fig , 8 View Fig E-F, 9A). Otherwise, the head is filled with numerous bubble-shaped structures of apparently undifferentiated cells. The inner armature in the mouth cone appears to consist of six longitudinal stylets. The stylets are enwrapped by an apical ring at the tip of the mouth cone, and by another posterior ring at the transition to the pharyngeal bulb. Small placoid-like plates attach posterior to the apical ring. The pharyngeal bulb is perfectly globular, but it does not seem to contain any differentiated structures, or any indication of cross-striated musculature. The ring-shaped brain is located apically in the head, and the inner mouth cone armature is passing through the ring ( Fig. 6B View Fig ). The brain consists of numerous, densely packed cells. It was possible to identify four pairs of muscles passing through the head: a dorsal pair, a laterodorsal, and lateroventral and a ventral pair. All muscles attach anteriorly near the mouth cone. Posteriorly, they appear to continue into the densely packed musculature of the trunk.
The trunk appears like a pedicle posterior to the large head. The length is about 70 μm, and it is 26 μm wide anteriorly and 20 μm wide in its slightly narrower posterior end. Internally the trunk is dominated by muscles ( Figs. 6B View Fig , 8B View Fig and 9A View Fig ). Numerous (+25) longitudinal muscles stretch through the trunk, from the posterior end and towards to the transition between trunk and head, where most of them have their anterior attachment point. Only four pairs continues through the head (see description above) and attach near the mouth cone. The longitudinal muscles are very thin (ca 0.8 μm) in the posterior end, and expand and broaden (ca 2 μm) as they progress towards the head. About twelve circular muscles wrap around the longitudinal ones. A single circular muscle is located close to the posterior end, and another double set of circulars is also fairly close to the hind end, but besides these, all other circular muscles are concentrated in the anterior 2/3 of the trunk. The circular muscles are generally thicker than the longitudinal ones, about 2–3 μm in width.
3.2.8. Notes on diagnostic features in Patuloricus tangaroa gen . et sp. nov
The distinct larval shape of P. tangaroa gen. et sp. nov. differs from most other Higgins larva of the three loriciferan families, and it solely shows resemblance to the only other known Shira larva - the larva of Tenuiloricus shirayamai Neves & Kristensen, 2014 . The two species (and genera) resemble each other by their general habitus, with a large bulbous head being wider than the trunk, and by the relatively long and slender thorax and abdomen. Additional similarities include the presence of six scalid rows in the introvert, the morphology of the clavoscalids that is nearly identical in the two species, and finally the accordion-like anatomy of the anterior part of the trunk. Neves & Kristensen (2014) use the level of the anteroventral setae to define the transition from thorax to abdomen in T. shirayamai . However, we prefer to see the entire accordion-like part of the trunk in P. tangaroa gen. et sp. nov. as an anatomical unit corresponding to the thorax. Likewise, we also interpret the thorax of T. shirayamai as representing a much larger portion of the trunk. With this interpretation, we have another similarity between the two species, namely anterior setae attaching 1/3 to halfway up the thorax, rather than in the transition between thorax and abdomen. We are aware that the anterior setae traditionally have been used to mark, perhaps even define, the transition from thorax and abdomen, but we see the anatomical differences between the accordion-like thorax and the deeply plicated abdomen as more distinctive than these setal attachment sites. It should furthermore be noted that there are previous reports of Higgins larvae, e.g., P. diva , Urnaloricus gadi Heiner & Kristensen, 2009 , and Titaniloricus inexpectatovus Gad, 2005 , as well as P. apteryx sp. nov., with anterior setae clearly attaching on the thorax, rather than in the transition line (see Gad 2005a 2009a; Heiner & Kristensen 2009; present contribution). Finally, the two species share the presence of posterodorsal and posterolateral setae, and the lack of terminal setae.
The numerous similarities between the larvae of T. shirayamai and P. tangaroa gen. et sp. nov. clearly justify that both larvae are considered as Shira larvae – a special subcategory of Higgins larvae. However, we also see several differences between the two entities, which justifies that they are considered as two distinct genera. Generic differences include presence of a differentiated neck region in Tenuiloricus , the presence of two pairs of anterior setae in Patuloricus gen. nov. opposite to only one pair in Tenuiloricus , the differentiation of three abdominal regions in Tenuiloricus opposite to only one in Patuloricus gen. nov., and finally the difference in toe morphology, i.e., long acicular toes in Tenuiloricus opposite to abruptly tapered toes with offset tips in Patuloricus gen. nov. Additional differences are also expressed in the morphology of Row 2 to 6 scalids, and in the shapes and sizes of anterior and posterior setae, but these are differences we would expect to be more relevant in species differentiation.
3.2.9. Notes on postlarval morphology of Patuloricus tangaroa gen . et sp. nov
The description of T. shirayamai only includes morphological information about Shira larval morphology. Thus, with a different larva present inside the Shira larva of P. tangaroa gen. et sp. nov. we get, for the first time, a glimpse of the stage following a Shira larva. Due to this stage’ s lack of reproductive organs, we interpret it as a postlarva, rather than an adult. The stage leaves us with more questions than answers though. The complete lack of outer structures, in particular scalids or any other kind of introvert appendages, suggests that the postlarva is not free-living, but a stage destined to remain inside the cuticle of the Shira larva. The presence of postlarvae never leaving the Higgins larva has been documented from other species, such as Rugiloricus manuelae Pardos & Kristensen, 2013 (see Pardos & Kristensen 2013). However, this indication is strongly contradicted by the highly muscular trunk, which clearly suggests that the postlarva is a motile stage. This is supported by the presence of differentiated and well-developed inner armature in the mouth cone, suggesting that the postlarva is a stage that feeds. Thus, for now, the fate of the Shira larva remains enigmatic, but we see the finding of its postlarva as one step towards understanding its life cycle.
| NIWA |
National Institute of Water and Atmospheric Research |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.