
Parotoplana pythagorae, Delogu, Valentina & Curini-Galletti, Marco, 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.177648 |
|
DOI |
https://doi.org/10.5281/zenodo.6242258 |
|
persistent identifier |
https://treatment.plazi.org/id/039FC129-1E04-FFE6-FF61-6AFC88CCF967 |
|
treatment provided by |
Plazi |
|
scientific name |
Parotoplana pythagorae |
| status |
sp. nov. |
Parotoplana pythagorae sp. n.
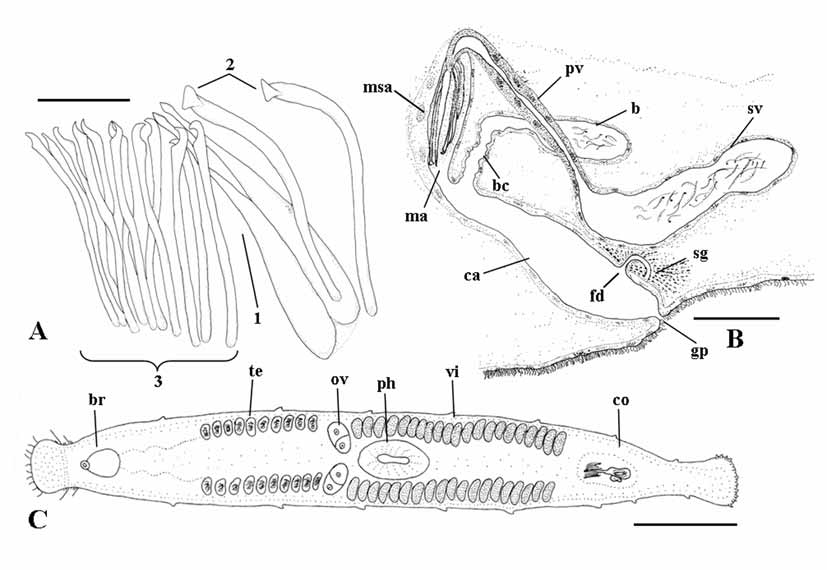
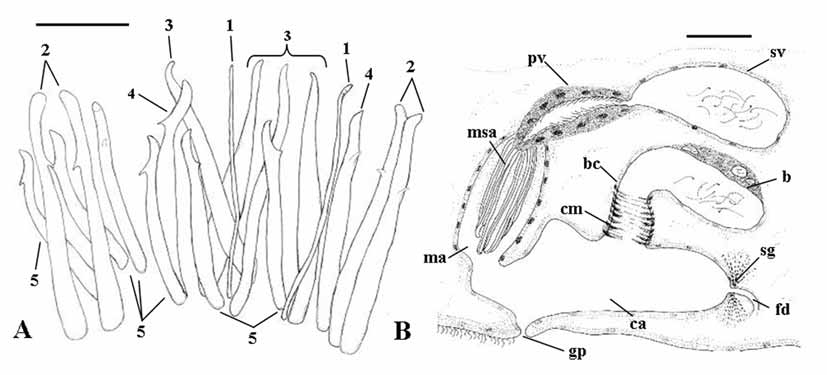
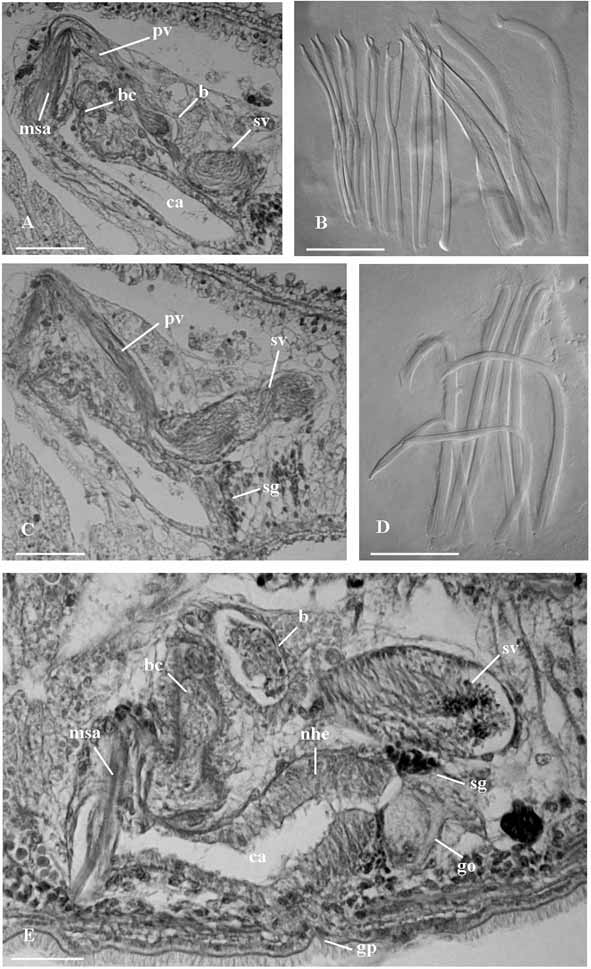
( Figs. 1 View FIGURE 1 A–C; Figs. 6 View FIGURE 6 A–C)
Holotype: one whole mount ( SMNH 6661).
Type locality: Apulia, Italy: Porto Cesareo (Lecce), Bacino Grande Bay, loc. Scala di Furno (lat. 40°16’26”N, long. 17°52’53.68”E), slightly silty coarse sand among rocks, lower intertidal in the Otoplana - zone. May 2005.
Additional material: Paratype ( SMNH 6662): one specimen from the type locality, sagittally sectioned. Six specimens sagittally sectioned (CZM-34/39); two karyological slides (CZM-40,41); two whole mounts (CZM-42,43), all from the type locality.
Etymology: the species name relates to the presence, in the sclerotized structures, of peculiar right-angled triangles, to which Pythagoras’ (Samos, 571-496 B.C.) most famous theorem refers.
Description. Living specimens agile, colourless and semi-transparent, of average size for the genus: the holotype, an adult worm, is about 1.3 mm long in fixed condition. The anterior end is clearly marked and provided with sensory bristles, and, ventrally, with a few adhesive papillae. The posterior end is fan-shaped and provided with very numerous adhesive papillae ( Fig. 1 View FIGURE 1 C). The encapsulated, oval shaped brain (about 60 µm long) abuts on a statocyst (about 21 μm in diameter).
Epithelium with intraepithelial nuclei. Ciliation restricted to a sensorial groove in the anterior end, and, ventrally, to a creeping sole that extends from the anterior end to posterior to the genital pore. The ciliated epithelium is markedly higher than the non-ciliated epithelium. Cilia up to 7 μm in length. Rhabdoids (up to 15 µm in length) are numerous dorsally and arranged in longitudinal rows.
The gut extends posteriorly nearly to the caudal end and anteriorly to the cephalic area. The nearly median pharynx is collar-shaped; its nucleated epithelium is ciliated except for a small area at the distal tip. Pharyngeal cilia are about 3 µm long. No esophageal area could be seen. The pharyngeal cavity opens to the outside through the mouth, which is surrounded by a ring of circular muscles.
Male genital organs. Testes, located anterior to the ovaries, consist of two lateral rows of about ten follicles each ( Fig. 1 View FIGURE 1 C). The copulatory organ consists of a sacciform seminal vesicle (about 105 μm long), connected distally to a prostatic vesicle ( Figs. 1 View FIGURE 1 B; 6 A, C). The seminal vesicle is lined by a nucleated epithelium. The prostatic vesicle is narrowly elongate and its nucleated epithelium is non ciliated. It is distally connected to the sclerotized apparatus ( Figs. 1 View FIGURE 1 A; 6 B) which consists of a central stylet (1), two companion spines (2) and a girdle of smaller spines (3). The stylet (95 μm long in the holotype, range 93–95 μm in the sample) wraps around the distal portion of the ejaculatory duct. In well-squeezed specimens, it is evident that the seemingly tubiform structure seen in semi-squashed living specimens consists of two spines connected for most of their length by a feebly sclerotized lamina. These spines are straight and their distal end is acutely rhomboidal. Two companion spines are placed laterally to the stylet. These spines are 96–98 µm long, slightly wider than the other spines, with distinctly triangular apices. Stylet and companion spines are surrounded by a girdle of 12 slender spines (69 to 76 µm long) with sickle shaped apices, provided with a somewhat obtuse subterminal tooth. The sclerotized apparatus is located within the male antrum, lined by a nucleated epithelium, which opens into the anterior portion of the common atrium.
Female genital organs. Two ovaries ( Fig. 1 View FIGURE 1 C) are located anterior to the pharynx. Two rows of yolk follicles (vitellaria) are present posterior to the ovaries to the level of the copulatory organ. In the sectioned material, it is possible to trace the presence of the female common duct only in its distal portion, at its opening into the common atrium, where it is surrounded by numerous shell glands ( Fig. 1 View FIGURE 1 B). The bursa ( Figs. 1 View FIGURE 1 B; 6 A), which is comparatively large and obvious in living specimens, opens into the common atrium close to the opening of the male system, by means of a wide canal about 70 µm long in the sectioned paratype, lined by an epithelium which appears irregular in shape, with numerous folds. The bursa, which is filled with sperm in all specimens sectioned, is lined by a low, nucleated epithelium. Male antrum, female duct and bursal canal open into a vast common genital atrium, which is lined by non ciliated, nucleated epithelium, surrounded by well developed longitudinal muscles. It opens to the outside through the common genital pore.
Karyotype. Chromosome number: n = 6; FN = 12. All chromosomes are isobrachial; the first two pairs are more than twice the size of the remaining chromosomes (Fig. 8 B). Chrom. I = r.l.: 28.49 + 0.68; c.i.: 46.06 + 0.55 (m); Chrom. II = r.l.: 27.80 + 0.21; c.i.: 44.04 + 0.66 (m); Chrom. III = r.l.: 12.3 + 0.59; c.i.: 35.8 + 3.01 (sm); Chrom. IV = r.l.: 11.99 + 0.76; c.i.: 46.73 + 0.49 (m); Chrom. V = r.l.: 9.94 + 0.18; c.i.: 30.06 + 0.7 (sm); Chrom. VI = r.l.: 9.46 + 0.46; c.i.: 36.86 + 2.66 (sm) (based on the measurements of five spermatogonial plates).
Remarks. Among the species of Parotoplana provided with a stylet (P. re n a t a e; P. papii Ax 1956 ; P. procerostyla Ax 1956 ; P. pacifica Ax & Ax 1967 ; P. macrostyla Lanfranchi 1978 and P. uncinata Lanfranchi 1978 ) only the Mediterranean P. renatae and P. macrostyla share with P. pythagorae sp. n. a general arrangement of spination consisting of a central stylet, two flanking companion spines and a girdle of smaller spines. Size of the sclerotized structures is comparable: the stylet is about 84–104 μm in P. renatae , 87–90 μm in P. macrostyla and 93–95 μm in the new species; companion spines reach respectively 100 μm (P. re n a t a e), 87– 90 μm ( P. macrostyla ) and 96–98 μm ( P. pythagorae sp.n.) in length; girdle spines are about 71–77 μm long in P. renatae , 74–77 μm in P. macrostyla and 69 to 76 μm in P. pythagorae sp.n. However, in none of the aforementioned species, are the companion spines provided with a distinctly triangular apex, easily appreciable even in semi-squashed living specimens. On the contrary, the apex of the companion spines is sickle-shaped in P. macrostyla and P. renatae ( Ax, 1956, Fig. 158, pg. 225), which is provided with a triangular, laminar stylet. The stylet of P. macrostyla has a split apex ( Lanfranchi, 1978, Fig.1 View FIGURE 1 B, pg. 250), and thus somewhat similar to the new species. However, in P. macrostyla the common female duct is virtually not existing, and the two oviducts join just at their opening into the common atrium ( Lanfranchi, 1978, Fig. 3 View FIGURE 3 , pg. 250). On the contrary, the existence of a common female duct could be observed in all the sectioned specimens of P. pythagorae sp. n.
The only species of Parotoplana whose karyotype is known is P. m a c ro s t y l a. The karyotype of P. pythagorae sp. n. appears very similar for number and relative size of chromosomes. However, in this species all chromosomes are isobrachial, whereas in P. macrostyla Chrom. VI is subtelocentric, with low centromeric index ( Curini-Galletti et al., 1984).
Diagnosis: Species of Parotoplana with a wide bursal canal, lined with a corrugated epithelium. The sclerotized apparatus of the copulatory organ consists of a stylet (93–95 μm long); two companion spines with triangular apex (96–98 μm long) and a girdle of 12 spines, provided with sickle shaped apices and subterminal, obtuse tooth (69–76 μm long). Karyotype with n = 6, and all chromosomes isobrachial.
| SMNH |
Saskatchewan Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |