
Parkia lutea H.C. Hopkins, 2000
|
publication ID |
https://doi.org/ 10.5281/zenodo.4605724 |
|
persistent identifier |
https://treatment.plazi.org/id/03CC9B7A-E532-FFE5-3734-FDAAAF946073 |
|
treatment provided by |
Carolina |
|
scientific name |
Parkia lutea H.C. Hopkins |
| status |
sp. nov. |
Parkia lutea H.C. Hopkins , sp. nov.
Parkia igneiflorae Ducke arcte affinis, sed oblongis majoribus (folii maximo foliolo 4.5-5.1 × 2-2.2 cm, non 2.7-4 × 0.55-1.2 cm) foliolis, paucipinnatis (2-3 non 3-7[-10]) et paucijugatis (8-11 non 17-26) foliis, praecipue differt.
TYPUS. — Zarucchi et al. 3219, Brazil, Amazonas ( AM) , Mun. de Maués, along Rio Apoquitaua above confluence of Rio Pacoval , 3º54’S, 57º53’W, fl., 27 July 1983 (holo-, GoogleMaps INPA [no. 154473]; iso-, K!, NY!, US!, perhaps others?) .
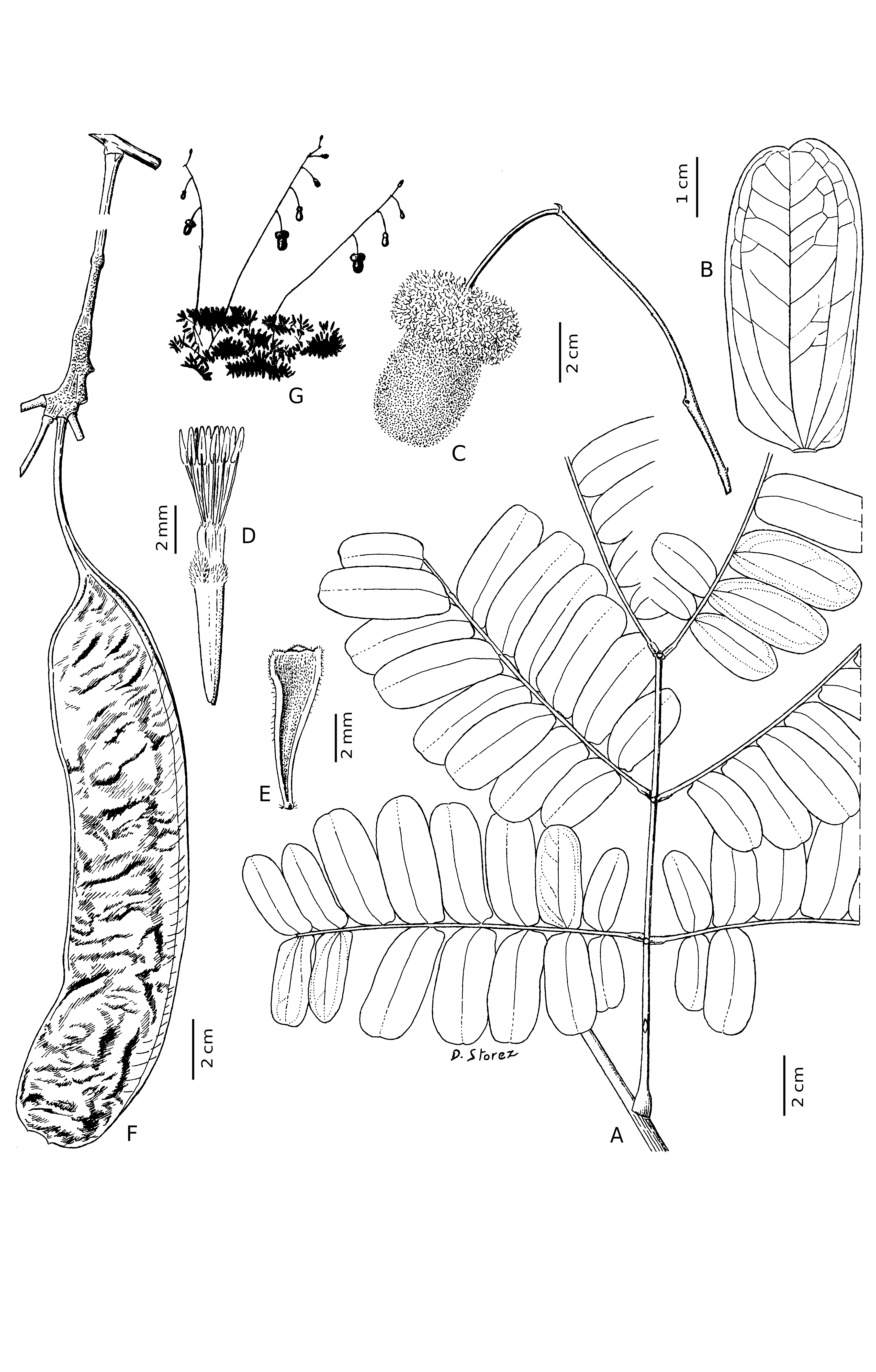
Small tree 5-10 m high, 15-25 cm dbh, the crown spreading or umbrella-shaped. Leaves bipinnate, alternate, sometimes widely spaced on stems, up to 24 cm apart. Leaf rachis 10.5-15.5 cm long, including petiole of 3.3-6 cm, with some minute indumentum, glabrescent; gland on adaxial surface of petiole ca. 1-2 cm above pulvinus, elliptical; glands on leaf rachis between points of attachment of opposite pinnae, round. Pinnae 2-3 pairs per leaf, up to 16.5 cm long; rachis of pinna with some minute indumentum, glabrescent. Leaflets 8-11 pairs per pinna, opposite or the most proximal pair sometimes subopposite, oblong, the base truncate, the apex rounded, broadly obtuse to truncate or emarginate, sides of leaflets parallel or leaflet sometimes slightly wider at the base; largest leaflet per leaf 4.5-5.1 × 2-2.2 cm; central vein placed somewhat towards distal margin, straight at apex and curved towards proximal side at base; secondary veins arising from point of attachment of leaflet to rachis of pinna; tertiary venation scarcely visible above but minutely prominent below; leaflets drying dark brown and shiny above, dull chestnut-brown below, glabrous, without exfoliating waxy layer.
Inflorescence axis woody, ascending, more than 26 cm long, bearing several alternate peduncles ca. 3-16.5 cm apart; inflorescence axis and peduncles bearing short, red-brown indumentum, glabrescent. Peduncles somewhat woody, 4- 5 (- 6 in fruit) cm long, 2 (- 5 in fruit) mm diameter, pendent. Capitula in bud brown, velutinous from indumentum on apices of imbricate floral bracts and calyx lobes. Capitula at anthesis biglobose, when dry ca. 6.2 cm long, the basal staminodial fringe 4.5 (or more) cm diameter, the constricted ring above fringe ca. 2.2 cm diameter, the apical ball of fertile flowers ellipsoid, ca. 3 cm diameter; staminodial fringe and fertile flowers egg-yolk yellow. Flowers small, tubular, very numerous and densely packed, inserted on a swollen, central axis, each subtend- ed by a small bract, of several functional types in each capitulum. Floral bracts up to 6.6 mm long, narrowly triangular, 0.3 mm wide at base and up to 2.1 mm wide at thickened, pubescent distal end. Fertile flowers forming a distal ball; calyx ca. 5.8 mm long including pseudopedicel ca. 0.5 mm long, tubular below with 5 unequal minutely pubescent lobes at distal end; corolla membranous, tubular with 5 short apical lobes ca. 0.6 mm long, tending to split at base into 5 segments (see LUCKOW & HOPKINS 1995, fig. 2b); filaments 10, united into a tube to approximately the level of the mouth of the calyx then free above for 4 mm; above pseudopedicel, corolla and filament tube united at base for ca. 3 mm, then free above; anthers ca. 1 mm long; gynoecium rudimentary. Staminodial flowers around base of capitulum: calyx 6-6.4 mm long including pseudopedicel, 0.5-0.8 mm diameter at the base, united below into a tube with 5 unequal, minutely pubescent lobes at distal end; corolla membranous, tubular, with 5 apical lobes exserted ca. 0.6-1.5 mm beyond mouth of calyx; filaments 10, exserted 14-20 mm beyond mouth of calyx, at least sometimes irregularly united into a tube towards the base, free above, somewhat crispate.
Fruit a flattened, indehiscent pod, several (ca. 7) together arising from swollen axis of capitulum, pendent; stipe ca. 4.5-5 cm long, valves 12- 19.5 × 4.2-4.4 cm, oblong or falcate, surface dark brown, minutely cracked, without indumentum, the base attenuate into stipe and the apex rounded; suture to which funicles attached narrow, the opposite suture flattened, ca. 6 mm wide; endocarp yellow, papery; seeds black, ca. 13-15 per pod. — Figs. 1 View Fig , 2. View Fig
ECOLOGY AND DISTRIBUTION. — Occurs in low forest with ajuru palms (name not traced) (Froés 33631) and in open white-sand campina, sometimes at the high water mark along the edge of igarapés (Zarucchi 3219). Flowering is recorded in July and fruits were collected in July and October. Known from a small area near the Rio Abacaxis and Rio Apoquitaua in Amazonas, Brazil ( Fig. 3 View Fig ).
PARATYPES. — BRAZIL: Froés 33631 , AM, região de Rio Abacaxis , Rio Marimarí, fr., 21 Oct. 1957 ( IAN! × 2); Zarucchi et al. 2930 , AM, Mun. de Axinim , basin of Rio Abacaxis , lower Rio Paca ca. 1 km from its confluence with the Rio Marimarí, 4º07’S, 58º58’W, fr., 1 July 1983 ( INPA [no. 154183], K!, NY!, US!); Zarucchi et al. 2945 GoogleMaps , AM, Mun. de Axinim , basin of Rio Abacaxis, ca. 4 km from mouth of Igarapé Surubim, 4º31’S, 58º34’W, fl., 4 July 1983 ( INPA [no. 154199], K!, NY!, US!) GoogleMaps .
DISCUSSION. — The description is based primarily on the type collection with the characters of the pods taken from Froés 33631 and Zarucchi 2930. The specific epithet refers to the colour of the capitula.
The shape and structure of the capitula (biglobose, with a well developed staminodial fringe at the base and a distal ball of fertile flowers) and the pods (indehiscent with a broad, flattened suture on the opposite side to the funicular attachment) indicate that this species belongs to the neotropical “nitida group” of Parkia sect. Parkia ( LUCKOW & HOPKINS 1995) . Most members of this group are known to be pollinated by phyllostomid bats ( DE CARVALHO 1960; HOPKINS 1984; VOGEL 1968) and the structure of the inflorescence and capitula suggest that P. lutea is also chiropterophilous.
The rather distinctive inflorescence, in which a somewhat woody, erect axis projects above the foliage and bears several pendent capitula, each on a long, flexible peduncle ( Figs. 1G View Fig , 2 View Fig A-C), occurs in several species of this group, and is especially similar to the inflorescence in P. igneiflora ( HOPKINS 1986, figs. 3A, 20A). As in other species of Parkia sect. Parkia , the basal staminodial flowers start to open before the fertile ones, which are in a distal ball ( Figs. 2 View Fig C-D). In P. lutea the staminodial and fertile flowers are of the same colour and shade while in some other neotropical species of Parkia sect. Parkia with yellow fertile flowers and a well-developed fringe, the staminodia are red ( P. panurensis Benth. ex H.C. Hopkins ) or white to cream (e.g. P. nitida Miq. , P. truncata Cowan ) ( HOPKINS 1984, 1986).
The only capitulum on the sheets to hand with flowers at anthesis has fertile flowers that are functionally male, since each has a small, rudimentary gynoecium. When considering the sexual expression of the flowers in the distal, fertile ball only, and discounting the specialised nectariferous and staminodial flowers, andromonoecy is common in Parkia and probably also occurs here.
It was not possible to verify the presence of nectariferous flowers in the narrow zone between the ball of fertile flowers and the staminodial fringe without destroying the entire capitulum, but such flowers are present in all other species of sect. Parkia . They differ from the fertile flowers in having a much thickened base, the filaments are not exserted, and the gynoecium is usually absent. Diagnoses for the three sections of Parkia are given in HOPKINS (1986).
Parkia lutea is closely related to P. igneiflora but differs in having leaves with fewer pairs of pinnae and fewer, larger leaflets, and pods which are generally smaller and rather similar to those of P. discolor Spruce ex Benth. ; however, there is no discontinuity in dimensions between the pods of P. igneiflora and P. lutea . Most collections of P. igneiflora have reddish or purplish flower-heads ( HOPKINS 1986), although in some collections they are golden yellow and thus similar to P. lutea . These latter collections of P. igneiflora were treated by DUCKE (1938) as a distinct vari- ety, var. aurea , but their leaves cannot be distinguished from those of the red- and purple-flowered “varieties” of P. igneiflora . Parkia igneiflora and P. lutea both occur in low forest on sandy soil.
The only other species of neotropical Parkia with leaflets of a similar size is P. gigantocarpa Ducke. However , this is a large to very large forest tree with much larger capitula (15-21.5 cm long), longer pods (ca. 52-62 cm including the stipe), and the leaflets are acute at the apex with a white waxy deposit on the underside ( HOPKINS 1986).


| AM |
Australian Museum |
| INPA |
Instituto Nacional de Pesquisas da Amazonia |
| IAN |
Embrapa Amazônia Oriental |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Parkia lutea H.C. Hopkins
| Hopkins, Helen C. F. 2000 |
Parkia lutea
| H. C. Hopkins 2000 |
P. lutea
| H. C. Hopkins 2000 |
P. lutea
| H. C. Hopkins 2000 |
P. lutea
| H. C. Hopkins 2000 |
P. igneiflora
| Ducke 1932 |
P. igneiflora
| Ducke 1932 |
P. igneiflora
| Ducke 1932 |
P. igneiflora
| Ducke 1932 |
P. igneiflora
| Ducke 1932 |
Parkia igneiflora
| Ducke 1932 |
P. gigantocarpa
| Ducke. However 1915 |