
Palaemon septemtrionalis, Katogi & Chiba & Yokoyama & Hatakeyama & Shirai & Komai, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4576.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:AD31C9CB-0470-4625-A56E-0363A60673CA |
|
DOI |
https://doi.org/10.5281/zenodo.5942624 |
|
persistent identifier |
https://treatment.plazi.org/id/03B387F9-FF8F-FF9D-A18F-FEAAFD92F876 |
|
treatment provided by |
Plazi |
|
scientific name |
Palaemon septemtrionalis |
| status |
sp. nov. |
Palaemon septemtrionalis View in CoL n. sp.
[New Japanese name: Kitano-suji-ebi]
Figs 1–7 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7
Material examined. Holotype: CBM-ZC 14920 , male (cl 10.9 mm), Isatomae River, Utatsu, Minami-sanriku Town , Miyagi Prefecture, 38°43’52.82”N, 141°28’09.60”E, upstream, mixture of sand, gravel and rock, among roots of reed, 23 February 2018, coll. Y. Katogi. GoogleMaps
Paratypes: CBM-ZC 14921 , 1 male (cl 11.3 mm), Isatomae River, Utatsu, Minami-sanriku Town , Miyagi Prefecture, 25 July 2017, coll. Y. Katogi ; OUMNH ZC 2018-01 - 074 , 1 male (cl 11.3 mm), same data ; CBM-ZC 14922 , 1 male (cl 12.5 mm), same locality, 10 May 2017, coll. Y. Katogi ; CBM-ZC 14923 , 1 male (cl 10.5 mm), same data as holotype GoogleMaps ; CBM-ZC 14924 , 1 male (cl 10.4 mm), same data as holotype GoogleMaps ; CBM-ZC 14925 , 1 female (cl 14.7 mm), same data as holotype GoogleMaps ; OUMNH ZC 2018-01 - 075 , 1 female (cl 11.7 mm), same data as holotype; CBM- GoogleMaps ZC 14926 , 1 male (cl 12.8 mm), Nishi-moune River, Karakuwa, Kesennuma City , Miyagi Prefecture, 38°54’11.85”N, 141°36’58.05”E, mid-to upstream, mixture of sand, gravel and rock, 25 February 2017, coll. Y. Katogi GoogleMaps ; CBM-ZC 14927 , 1 male (cl 14.8 mm), same locality, 11 May 2017, coll. Y. Katogi GoogleMaps ; CBM-ZC 14928 , 1 male (cl 14.0 mm), same locality, 15 November 2017, coll. Y. Katogi. GoogleMaps
Other material: NSMT-Cr 1541, 1 ovigerous female (cl 12.7 mm), Kita-naganuma Pond, creek of east side, Sendai City, Miyagi Prefecture, 20 October 1974; NSMT-Cr 1543, 1 female (cl 16.4 mm), 1 ovigerous female (cl 13.9 mm), junction of Natori River and Koishi River, Sendai City , Miyagi Prefecture, 28 July 1974 ; NSMT-Cr 1691, 1 female (cl 15.1 mm), junction of Hirose River and Natori River, Sendai City , Miyagi Prefecture, 26 July 1974, coll. M. Nakamura ; NSMT-Cr 1703, 1 ovigerous female (cl 14.8 mm), Natori River, Sendai City , Miyagi Prefecture, 16 June 1974, coll. M. Nakamura ; NSMT-Cr 20017, 1 female (cl 13.3 mm), Bagyuu-numa Pond, Katsuta District , Miyagi Prefecture, 15 November 1931 ; NSMT-Cr 20082, 2 females (cl 14.7 mm, 15.5 mm), Torinoumi, Watari Town , Miyagi Prefecture, 15 February 1933, coll. S. Atsumi ; NSMT-Cr 20095, 2 ovigerous females (cl 15.6 mm, 16.0 mm), Asamai, Hiraka Town, Yokote City , Akita Prefecture, 2 June 1932, coll. K. Ono.
Comparative material: Palaemon paucidens : NSMT-Cr 20038, 4 females (cl 12.6–14.1 mm), Okada, Miyagino-ku, Sendai City, Miyagi Prefecture, 17 October 1931; NSMT-Cr 20082, 9 females (cl 14.1–16.0 mm), Torinoumi, Watari Town, Sendai City, Miyagi Prefecture, 15 February 1933, coll. S. Atsumi; NSMT-Cr 1713, 1 male (cl 8.8 mm), 2 females (cl 10.3, 11.0 mm), Natori River, Sendai City, Miyagi Prefecture, 28 July 1975, coll. M. Nakamura; CBM-ZC 7829 , 1 female (cl 11.3 mm), Shido River, Inzai City , Chiba Prefecture, February 2004, coll. Y. Matsuzawa ; CBM-ZC 5165 , 1 female (cl 9.7 mm), Iinozawa, Sakura City , Chiba Prefecture, 29 July 1999, coll. H. Niijima ; CBM-ZC 8722 , 1 male (cl 13.3 mm), 1 ovigerous female (cl 8.7 mm), Kawajiri, Kamisu City, Ibaraki Prefecture, Tone River , 27 April 2006, coll. Y. Iga; Kawajiri, Tone River , Kamisu City , Ibaraki Prefecture, 27 April 2006, coll. T. Komai ; CBM-ZC 10466 , 4 males (cl 5.7–7.1 mm), 2 females (cl 9.0, 10.4 mm), junction of Takane River and Unaka River, Yasaka City , Kyoto Prefecture, 7 April 2007, coll. S. Saito ; CBM-ZC 14929 , 1 female (cl 10.8 mm), Nishi-moune River, Karakuwa, Kesennuma City , Miyagi Prefecture, 38°54’11.85”N, 141°36’58.05”E, mid-to upstream, mixture of sand, gravel and rock, 9 May 2017, dip net, coll. Y. Katogi; CBM- GoogleMaps ZC 14930 , 1 female (cl 9.7 mm), same data GoogleMaps ; CBM-ZC 14931 , 1 female (cl 9.2 mm), same data GoogleMaps ; CBM-ZC 14932 , 1 male (cl 9.3 mm, photo), same locality GoogleMaps , 17 June 2018, dip net, coll. Y. Katogi; CBM-ZC 14933 , 1 female (cl 9.5 mm), Tsukuba Botanical Garden, National Museum of Nature and Science , Tsukuba City , Ibaraki Prefecture, 36°06’04”N, 140°06’45”E, 26 September 2017, dip net, coll. Y. Katogi and T. Komai GoogleMaps ; CBM-ZC 14934 , 1 female (cl 9.6 mm), same data GoogleMaps .
Palaemon sinensis : CBM-ZC 13755 , 2 specimens, Enshu Beach, Hamamatsu City , Shizuoka Prefecture, 34°65.15’N, 137°77.87’E, freshwater pond, 7 January 2017, dip net, coll. D. Furukawa.
Diagnosis. Rostrum directed forward or slightly downward, faintly sinuous or nearly straight in lateral view, 0.6–0.8 times as long as carapace, slightly overreaching to falling slightly short of distal end of antennular peduncle; dorsal margin armed with 6–8 teeth, including 1 postorbital located at 0.1 of carapace length and 1 or 2 small subterminal; ventral margin with 2 or 3 teeth in distal half. Carapace smooth; branchiostegal spine subequal in size to antennal spine, placed on anterolateral margin of carapace; branchiostegal suture extending from just superior to base of branchiostegal spine to 0.4 of carapace length, slightly arcuate. Telson with posterior margin normally drawn out into small spine flanked by 2 unequal pairs of spiniform setae. Maximal diameter of cornea slightly less than 0.2 of carapace length. Pereopods 2 with chelae 1.1–1.4 times as long as carpus and about 5.0 times as long as wide; dactylus 0.7 times as long as palm. Pereopods 3–5 moderately stout; dactylus about 0.3 times as long as propodus in pereopod 3, flexor margin with low laminar convexity just proximal to base of unguis.
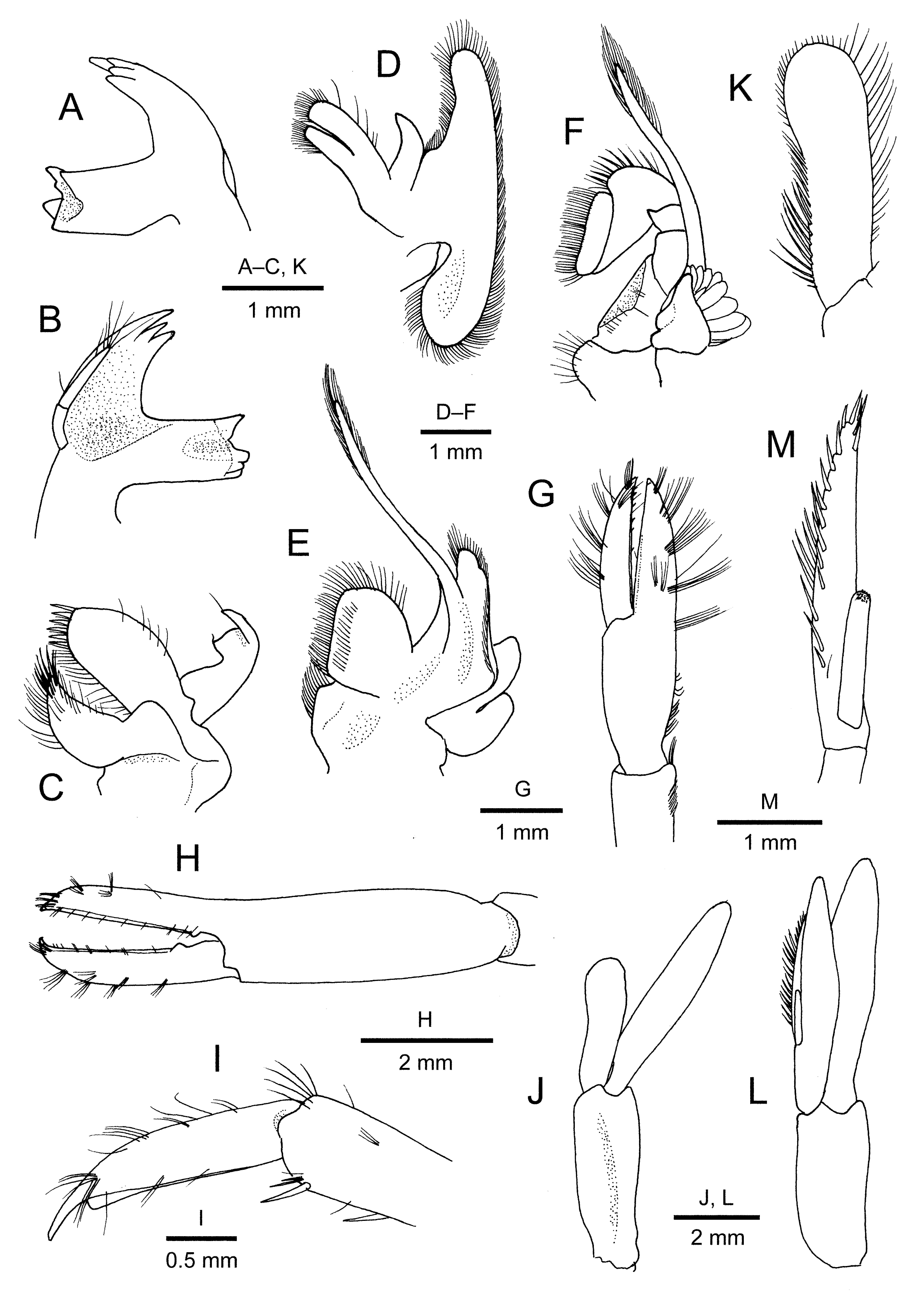
Description. Rostrum ( Figs 1A View FIGURE 1 ; 2A, B View FIGURE 2 ; 5A, B View FIGURE 5 ) directed forward or slightly downward, faintly sinuous or nearly straight in lateral view, 0.6–0.8 times as long as carapace, slightly overreaching or falling slightly short of distal end of antennular peduncle; dorsal margin armed with 6–8 teeth, including 1 postorbital located at 0.1 of carapace length and 1 or 2 small subterminal; short row of setae anterior to base of each tooth (except for subdistal tooth); ventral margin with 2 or 3 ventral teeth in distal half, and with 2 rows of short plumose setae interrupted at tip of each tooth; distal half of ventral margin forming shallow blade with gently convex lateral profile; lateral ridge obsolescent.
Carapace ( Figs 1A View FIGURE 1 ; 2A, B View FIGURE 2 ) smooth, glabrous; postrostral median ridge low, not extending to midlength of carapace; orbital margin evenly concave, inferiorly with rounded suborbital lobe; branchiostegal spine subequal in size to antennal spine, placed on concave anterolateral margin of carapace; branchiostegal suture extending from just superior to base of branchiostegal spine to 0.4 of carapace length, slightly curved.
Thoracic sternite 4 with small, anteriorly directed median spine on anterior margin, and with short transverse ridge medially widely interrupted on ventral surface. Sternite 5 with paired low tubercles on ventral surface. Sternites 6 and 7 without conspicuous ornamentation. Sternite 8 with small, anteroventrally directed median spine in males, only with low median protuberance in females.
Pleon ( Fig. 1B View FIGURE 1 ) of usual shape of Palaemon species. Pleomeres 1–4 with rounded pleura. Pleomere 5 with small posteroventral spine on pleuron. Sternites of pleomeres 1–4 each with median tubercle becoming smaller and less distinct posteriorly; sternite 5 with low, crest-like median ridge. Pleomere 6 twice as long as pleomere 5, 1.8 times as long as proximal depth, with small spine at posteroventral angle; posterolateral process subtriangular, terminating in small spine; sternite with small tubercle at each posterolateral angle; preanal area ( Fig. 2C View FIGURE 2 ) with sharp median carina.
Telson ( Figs 1B View FIGURE 1 ; 2D, E View FIGURE 2 ) 3.6 times as long as wide, with 2 pairs of dorsolateral spiniform setae, none marginal, anterior pair located at 0.5–0.6 of telson length and posterior pair at 0.7–0.8; lateral margin with row of very short setae (easily broken off in preservative); posterior margin normally drawn out into small spine, flanked and exceeded normally by 2 pairs of greatly unequal spiniform setae (mesial pair exceeding 3 times as long as lateral pair) (in holotype, one extra minute spiniform seta present on left side).
Eye (including cornea) ( Figs 1A View FIGURE 1 , 2B View FIGURE 2 ) subpyriform; cornea slightly wider than stalk, maximal diameter 0.18 of carapace length; small ocellar spot (= nebenaugen of Johnson et al. 2015) evident on dorsal surface.
Antennular peduncle ( Fig. 2B View FIGURE 2 ) not reaching distal margin of antennal scaphocerite. Article 1 with lateral side compressed dorsolaterally, with spiniform stylocerite reaching to 0.4 length of article; distolateral spine overreaching obliquely roundly truncate distal margin of lateral extension and overreaching midlength of peduncular article 2. Article 2 subequal in length to article 3, distolateral and distomesial margins of article 2 produced distally to accommodate basal part of article 3. Outer flagellum exceeding 1.5 times as long as carapace, bifurcate, fused portion consisting of about 10 articles ( Fig. 2F View FIGURE 2 ); shorter ramus less than half length of longer ramus, consisting of 30 or more articles, these articles each with concavity on lower lateral side. Inner flagellum thin, slightly shorter than outer flagellum.
Antennal peduncle ( Fig. 2G View FIGURE 2 ) with stout basicerite armed with moderately small ventrolateral distal spine. Scaphocerite 0.8 times as long as carapace, 3.2 times as long as wide; lateral margin nearly straight, but curving inward near base of distolateral spine; distolateral spine triangular, falling far short of rounded distal lamella. Flagellum exceeding 4 times as long as carapace.
Mouthparts ( Fig. 3 View FIGURE 3 A–F) typical of genus, as illustrated. Mandible always with 2-articulated palp.
Maxilliped 3 ( Fig. 4A View FIGURE 4 ) endopod nearly reaching to midlength of antennal scaphocerite. Coxa with subsemicircular lateral process. Antepenultimate article dorsoventrally flattened in proximal half, dorsally elevated in distal half. Penultimate article subcylindrical, with stiff setae on extensor surface mesially. Ultimate article 0.9 times as long as penultimate article, tapering to spiniform corneous tip; extensor margin to mesial face with numerous stiff setae consisting grooming apparatus.
Pereopod 1 ( Figs 3G View FIGURE 3 ; 4B View FIGURE 4 ) almost reaching to distal margin of antennal scaphocerite. Ischium shorter than merus; ventral margin slightly expanded, with several stiff setae. Merus 1.5 times as long as ischium, slightly compressed laterally. Carpus widened distally, about twice length of chela. Chela with grooming apparatus on mesial surface of palm; fingers with several tufts of short stiff setae on both lateral and mesial surfaces; occlusal margin of fixed finger forming thin corneous edge; dactylus subequal in length to palm, occlusal margin with row of minute spiniform setae.
Pereopod 2 ( Figs 3H View FIGURE 3 ; 4C View FIGURE 4 ) longest, slightly stouter than other pereopods, subequal in length and similar from right to left, overreaching antennal scale by length of chela. Ischium and merus slightly compressed laterally, subequal in length for each other, without armature. Carpus widened distally, 1.1 times as long as merus, about 5 times as long as distal width. Chela 1.1–1.4 times as long as carpus, about 5.0 times as long as wide, terminating in small corneous claw; fingers crossed at tips when closed; fixed finger proximally with small tooth on sharply edged occlusal margin; dactylus 0.7 times as long as palm, occlusal margin sharply edged, proximally with low convexity and small proximal tooth flanking occlusal tooth on fixed finger when closed.
Pereopod 3 ( Figs 3I View FIGURE 3 ; 4D View FIGURE 4 ) moderately stout, falling slightly short of distal margin of antennal scaphocerite. Ischium and merus slightly compressed laterally; merus about 2.6 times as long as ischium, unarmed. Carpus about half length of propodus, unarmed. Propodus with 2 longitudinal rows of evenly spaced, slender spiniform setae on flexor surface (5 or 6 in number in each row), extending onto terminal margin. Dactylus strongly compressed laterally, about 0.3 times as long as propodus, 4.6 times as long as basal height, gently curving distally, terminating in slender, corneous unguis; extensor margin with row of tufts of short, stiff setae; flexor margin bordered with thin, corneous ridge, forming low laminar convexity just proximal to unguis.
Pereopod 4 ( Fig. 4E View FIGURE 4 ) similar to pereopod 3.
Pereopod 5 ( Fig. 4F View FIGURE 4 ) generally similar to pereopod 3 and 4, but slightly longer, falling slightly short of distal margin of antennal scaphocerite. Propodus with grooming apparatus consisting of distolateral cluster of setae followed by 2 or 3 tufts of short stiff setae; flexor surface with 2 rows of small spiniform setae, lateral row with 5 or 6 spiniform setae proximal to grooming apparatus, mesial row with 10 or 11 spiniform setae extending onto terminal margin.
Male pleopod 1 ( Fig. 3J, K View FIGURE 3 ) with endopod reaching slightly beyond midlength of exopod, distal margin broadly rounded, mesial margin slightly sinuous but without notch, bearing row of bristle-like setae in proximal half. Male pleopod 2 ( Fig. 3L, M View FIGURE 3 ) with appendix interna less than half length of appendix masculina; appendix masculina elongate, reaching to distal 0.3 of endopod, dorsal margin slightly arcuate, ventral margin straight, tapering distally, armed with short to long, bristle-like setae or spiniform setae (about 20 in number) on dorsal margin to apex.
Uropod distinctly overreaching telson; exopod slightly longer than endopod, lateral margin terminating posteriorly in small spine far exceeded by posterior lamella, with small spiniform seta just mesial to posterolateral spine.
Variations. As is apparent from the above description, the direction and general shape of the rostrum is rather variable, as figured ( Fig. 1A View FIGURE 1 , 5A View FIGURE 5 ). One of the paratypes (CBM-ZC 14925, female, cl 14.7 mm) has an abnormal rostrum ( Fig. 5B View FIGURE 5 ), possibly linked to damage.
The telson is armed normally with two pairs of spiniform setae, but in one of the paratypes (CBM-ZC 14927, male, cl 14. 8 mm), there is only one spiniform seta on the right side. The length and number of spiniform setae on the distal end of the telson is substantially variable individually (see Figs. 2E View FIGURE 2 , 5 View FIGURE 5 D–G), although there are two pairs normally.
Coloration in life. Body ( Fig. 6A View FIGURE 6 ) and appendages generally light brown with numerous small brown chromatophores on semitransparent background. Carapace with pattern of dark brown stripes: one longitudinal stripe extending from orbital margin, but not reaching posterior margin of carapace; one short, obliquely longitudinal stripe extending to hepatic region; one transverse stripe across hepatic region; two obliquely transverse stripes on branchial region; one line running along anterolateral, ventral and posterolateral margin of carapace. Pleomeres 1–6 with posterior margins darker. Pereopod 1–5 each with yellowish band on distal part of ischium, merus, carpus and palm or propodus, tip of fingers or dactylus also yellowish; carpus and palm (pereopods 1 and 2) or propodus (pereopods 3–5) pale blue (except for yellow distal parts).
Etymology. From the Latin “ septemtrionalis ”, meaning “northern”, referring to the distribution of the new species, which is the northern part of the Japanese Archipelago.
Remarks. The present new species is morphologically very similar to P. paucidens in the general shape and armature of the rostrum and the general shape of the dactyli of the pereopods 3–5. The identity of Palaemon paucidens was confirmed through examination of the photograph of the lectotype presented by Yamaguchi & Baba (1993: fig 37), which clearly shows a slender pereopod 2 with the chela being shorter than carpus. Palaemon septemtrionalis n. sp. can be distinguished from P. paucidens by the structure of the pereopod 2 ( Fig. 4C View FIGURE 4 versus Fig. 7 View FIGURE 7 A–D). The chela of the pereopod 2 is longer than the carpus in the new species (chela length/carpus length 1.1–1.4), whereas the chela is usually shorter than the carpus in P. paucidens . Furthermore, the pereopod 2 is generally more robust in the new species than in P. paucidens . The dactylus of the pereopod 3 has a slight laminar convexity on the flexor margin just proximal to the unguis in P. septemtrionalis n. sp. ( Fig. 3I View FIGURE 3 ), but such a convexity is not seen in P. paucidens .
Palaemon sinensis View in CoL is also closely related genetically and morphologically to the new species and P. paucidens View in CoL (cf. Carvalho et al. 2017). The species was introduced from China and has been recorded from various Japanese localities (e.g., Oonuki et al. 2010; Imai & Oonuki 2014; Saito et al. 2017; Hiraoka et al. 2018). Nevertheless, P. septemtrionalis View in CoL n. sp. is easily distinguished from P. sinensis View in CoL by the branchiostegal suture originating from the base of the branchiostegal spine (versus originating superior to the branchiostegal spine in P. sinensis View in CoL ), the more robust pereopod 2 with the chela being longer than the carpus (versus shorter than the carpus) and the presence of the mandibular palp (absent in P. sinensis View in CoL ) ( Li et al. 2007; Imai & Oonuki 2014; this study). The size of the eye and the color pattern of the carapace, which are useful to differentiate between P. sinensis View in CoL and P. paucidens View in CoL (cf. Imai & Oonuki 2014), are also applicable in distinguishing between the new species and P. sinensis View in CoL .
Kubo (1938) indicated that Palaemon miyadii View in CoL , described on the basis of material from Manchoukuo, was closely allied to P. paucidens View in CoL , and thus it is worth to compare the new species with P. miyadii View in CoL . Comparison with the original description of Kubo (1938) confirms that the new taxon is not conspecific with P. miyadii View in CoL . The dorsal margin of the rostrum usually has a subterminal tooth in the new species, but there is no subterminal tooth in P. miyadii View in CoL . The appendix masculina of the male pleopod 2 of P. miyadii View in CoL is relatively shorter with less developed armature in P. miyadii View in CoL . Furthermore, from the description by Kubo, it can be assumed that the chela of the pereopod 2 is shorter than the carpus in P. miyadii View in CoL .
The phylogenetic analysis by Carvalho et al. (2017) suggested that Palaemon tonkinensis ( Sollaud, 1914) is in sister relation to P. paucidens . However, as the previous generic assignment of this species to Coutierella Sollaud, 1914 suggests, it is morphologically quite distinctive in Palaemon in the lack of a well-developed pleurobranch on the thoracomere 4, the presence of feebly developed, distinctly subspatulate chelae of the pereopod 2 and the presence of three or more pairs of spiniform setae on the posterior margin of the telson as well as several specialized features of the mouthparts ( Bruce 1989).
In the genetic analysis, a DNA matrix of 464 aligned base positions was obtained; 202 inserts and/or deletions were needed. The present specimens sequenced were composed of two and three haplotypes in Palaemon septemtrionalis n. sp. and P. paucidens , respectively. The unrooted NJ tree is shown in Fig. 8 View FIGURE 8 . The present new species shared an internal node (node A in Fig. 8 View FIGURE 8 ) only with P. paucidens (not with P. tonkinensis ), and their interspecific differences were genetically substantial (5.0–6.4% in p-distance). Node B was shared by four species, i.e. P. macrodactylus Rathbun, 1902 , P. gravieri ( Yu, 1930) , P. ogasawaraensis Kato & Takeda, 1981 , and P. serrifer ( Stimpson, 1860) , being similar with a terminal but distinct clade in Carvalho et al. (2017). These four species are genetically closely related for each other, and their interspecific divergence was 1.3–6.6% in p-distance. These results suggest that an appreciable period of time should have passed from divergence of P. septemtrionalis and P. paucidens , and thus these two taxa can be recognized as genetically independent species from each other.
On the basis of genetic analyses using the mitochondrial 16 rRNA gene, Chow et al. (2018b) recognized three genetically distinct lineages within P. paucidens they attributed to types A, B, and C. Specimens referred to the types A and B came from Hokkaido to Kyushu, but the type C is restricted to Amami-ohshima Island in the Ryukyu Islands. They also clarified that the type B shows regional genetic differentiation, divided into two major clades (subclades B1 and B2). Through a BLAST search on GenBank we found that 18 sequences of the 16 rRNA gene (Accession numbers LC384751 View Materials – LC384765 View Materials , LC384767 View Materials , LC384771 View Materials , LC384772 View Materials ), which constitute the subclade B1 of Chow et al. (2018b), match our new species with 99–100% agreement. The type A seems to represent P. paucidens , but the status of the subclade B2 and the type C needs to be assessed in the taxonomic scope. One of the authors (TK) personally examined specimens identified with the type A by Dr. Seinen Chow, confirming that those agree with P. paucidens as diagnosed in this study.
Distribution and habitat. During our field surveys, we have collected specimens of the new species from only two rivers in Miyagi Prefecture, Tohoku District, i.e., Nishi-moune River (Kesennuma City) and Isatomae River (Minami-sanriku Town). Specimens in the type series were collected from mid- to upstream of rivers; occurred on bottoms consisting of sand, gravel and rocks or among roots of reed. In these two rivers, Palaemon paucidens often coexisted with the new species.
Because of the close morphological similarity and the sympatric occurrence, it is expected that the new species might have been confounded with P. paucidens . Therefore, we examined material from various Japanese localities, identified with P. paucidens , in the collections of the NSMT and CBM (see “Comparative material”). Of the 451 specimens examined, 11 specimens from Akita and Miyagi Prefectures in the collection of NSMT were identified with the new species (see above “Other material”). Furthermore, sampling locations of 18 sequences of the 16S rRNA gene, representing clade B1 of P. paucidens , include Hokkaido, Aomori, Akita, Ishikawa, Kyoto, Hyogo and Miyagi Prefectures (cf. Chow et al. 2018: Appendix Table S1). In summary, the geographical range of the new species includes the Sea of Japan side ranging from Hokkaido to Hyogo Prefecture and the Pacific side ranging from Aomori to Miyagi Prefecture.







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Palaemon septemtrionalis
| Katogi, Yuichi, Chiba, Susumu, Yokoyama, Katsuhide, Hatakeyama, Makoto, Shirai, Shigeru & Komai, Tomoyuki 2019 |
P. septemtrionalis
| Katogi & Chiba & Yokoyama & Hatakeyama & Shirai & Komai 2019 |
P. paucidens
| De Haan 1844 |
P. paucidens
| De Haan 1844 |
P. paucidens
| De Haan 1844 |