
Pachnoda iskuulka Král, Sommer & Šípek, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4604.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:5068F1E3-0507-424D-8524-321BBAC1D58D |
|
persistent identifier |
https://treatment.plazi.org/id/03E9782F-716D-FFE1-36B5-C8F0FCB5198C |
|
treatment provided by |
Plazi |
|
scientific name |
Pachnoda iskuulka Král, Sommer & Šípek |
| status |
sp. nov. |
Pachnoda iskuulka Král, Sommer & Šípek , new species
( Figs. 1–9 View FIGURES 1–6 View FIGURES 7–12 , 19–20 View FIGURES 19, 20 , 41–42 View FIGURES 41–46 )
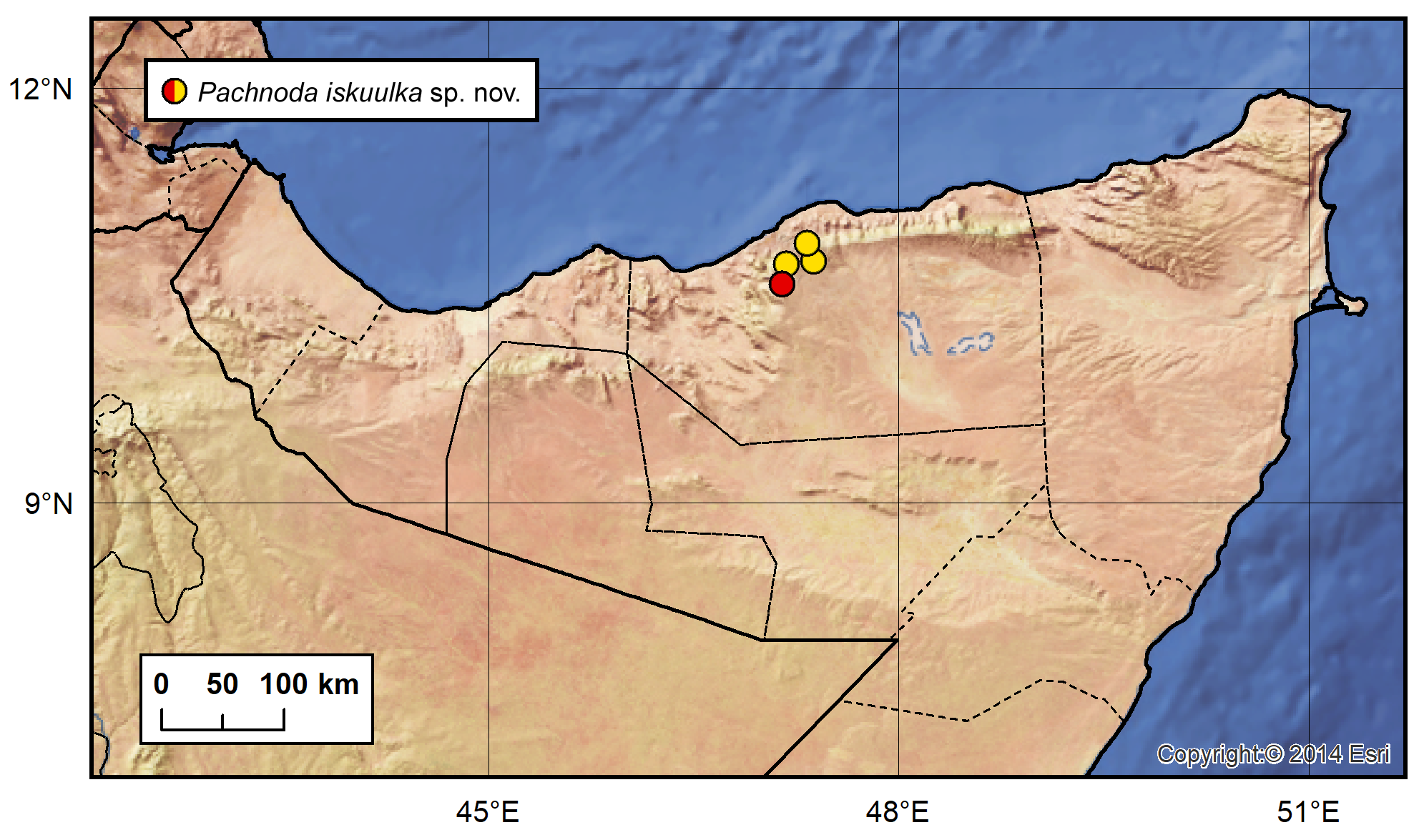
Type locality. Somaliland, Sanaag region, Buq village , surroundings of the school, 10°37’24”N 47°11’06”E, ca 1730 m a. s. l. GoogleMaps
Type material (43 specimens). Holotype, ♁ ( NMPC) , “Somaliland, 27.–28.viii.2018 | BUQ village - school | 10°37’24”N 47°11’06”E, ca 1730m | P. Kabátek, D. Král, D. Sommer lgt. [printed]” GoogleMaps . Paratypes: allotype, ♀ ( NMPC) , “Somaliland, 2.ix.2017 | DAALLO forest, view point | 10°46’14.01”N 47°17’52.07”E, ca 2150m | David Král lgt. [printed]”; paratypes, 1 GoogleMaps ♁ 1 ♀ ( PKCP) , 1 ♀ ( SJCP) [middle leg in pure alcohol in NMPC] “Somaliland, 22.–23.viii.2018 | DAALLO forest - base camp | 10°45’36”N 47°18’12”E, ca 2180m | P. Kabátek, D. Král, D. Som- mer lgt. [printed]”; 1 GoogleMaps ♁ 1 ♀ ( DSCS) “Somaliland, 23.–24.viii.2018 | W of MADERMOGE by road | 10°47’56”N 47°17’50”E, ca 1400 m | P. Kabátek, D. Král, D. Sommer lgt. [printed]”; 3 ♁♁ GoogleMaps 4 ♀♀ ( CUPC) , 6 ♁♁ 3 ♀♀ ( DSCS) , 1 ♁ (MNHN), 5 ♁♁ 3 ♀♀ ( PKCP) , 1 ♁ ( SJCP) [middle leg in pure alcohol in NMPC] “Somaliland, 27.–28.viii.2018 | BUQ village - school | 10°37’24”N 47°11’06”E, ca 1730m | P. Kabátek, D. Král, D. Sommer lgt. [printed]”; 2 ♁♁ GoogleMaps 2 ♀♀ ( CUPC) , 3 ♁♁ 2 ♀♀ ( DSCS) , 1♁ ( NMPC) “Somaliland, 27.–28.viii.2018 | BUQ village - school | 10°37’24”N 47°11’06”E, ca 1730m | P. Kabátek, D. Král, D. Sommer lgt. || F1 generation obtained | from wild-collected adults | date of emergence 20.i.2019 [printed]” GoogleMaps .
Description of holotype (♁) ( Figs. 1–3 View FIGURES 1–6 ). Body. Oval-shaped, rather flat, black, with white-orange-reddish pat- tern dorsally, slightly shining.
Dorsal surface. Head black; pronotum light orange, anterior margin black, base with narrow black strip project- ing in anvil-shaped spot medially; scutellum black; elytra black, with two orange spots just before middle almost fused with lateral longitudinal reddish spots, expanded posteriorly inside and connected to medial reddish spots, praeapical spots reddish and wide; pygidium with four small white spots along posterolateral margin ( Figs. 2–3 View FIGURES 1–6 ).
Ventral surface. Black, shiny; mesoventrum with white spot posterolaterally, metepiventrum with two small white spots, metepimeron with very small white longitudinal spot, joints of all legs with white spots ( Fig. 2 View FIGURES 1–6 ), each visible abdominal ventrite with small white strip laterally ( Fig. 2 View FIGURES 1–6 ).
Head. Clypeus bilobed; anteroclypeal margin mostly rounded; clypeolateral margins almost parallel; clypeolat- eral ridges rounded; disc prominently convex, clypeus anteriorly declivous; with coarse, scattered punctures.
Pronotum. Anterior pronotal margin concave or without prominence; lateral margins strongly converging ante- riad, regularly convex, lateral grooves absent; basolateral angles squared; basal margin strongly trisinuate, concave above elytral bases.
Scutellum. Slightly longer than wide; with apex rounded; with sides straight and entirely grooved.
Elytra with subhumeral arch rather shallow; preapical umbone prominent; with apicosutural angle mostly flat and rounded.
Pygidium with border thinly upturned; with scattered punctures and setae in transverse rows.
Ventrum ( Fig. 3 View FIGURES 1–6 ). Prosternum carinate medially. Meso-metaventral process broad; with sides concave; with an- terior margin convex; with mesoventral declivity concave. Abdominal ventrites with distinct longitudinal groove.
Legs ( Figs. 1–2 View FIGURES 1–6 ). Protibia tridentate. Meso- and metatibia with transverse carina on outside developed, slightly dentate; distal margin tridentate; tibial spurs unequal and metatibial spurs shortened.
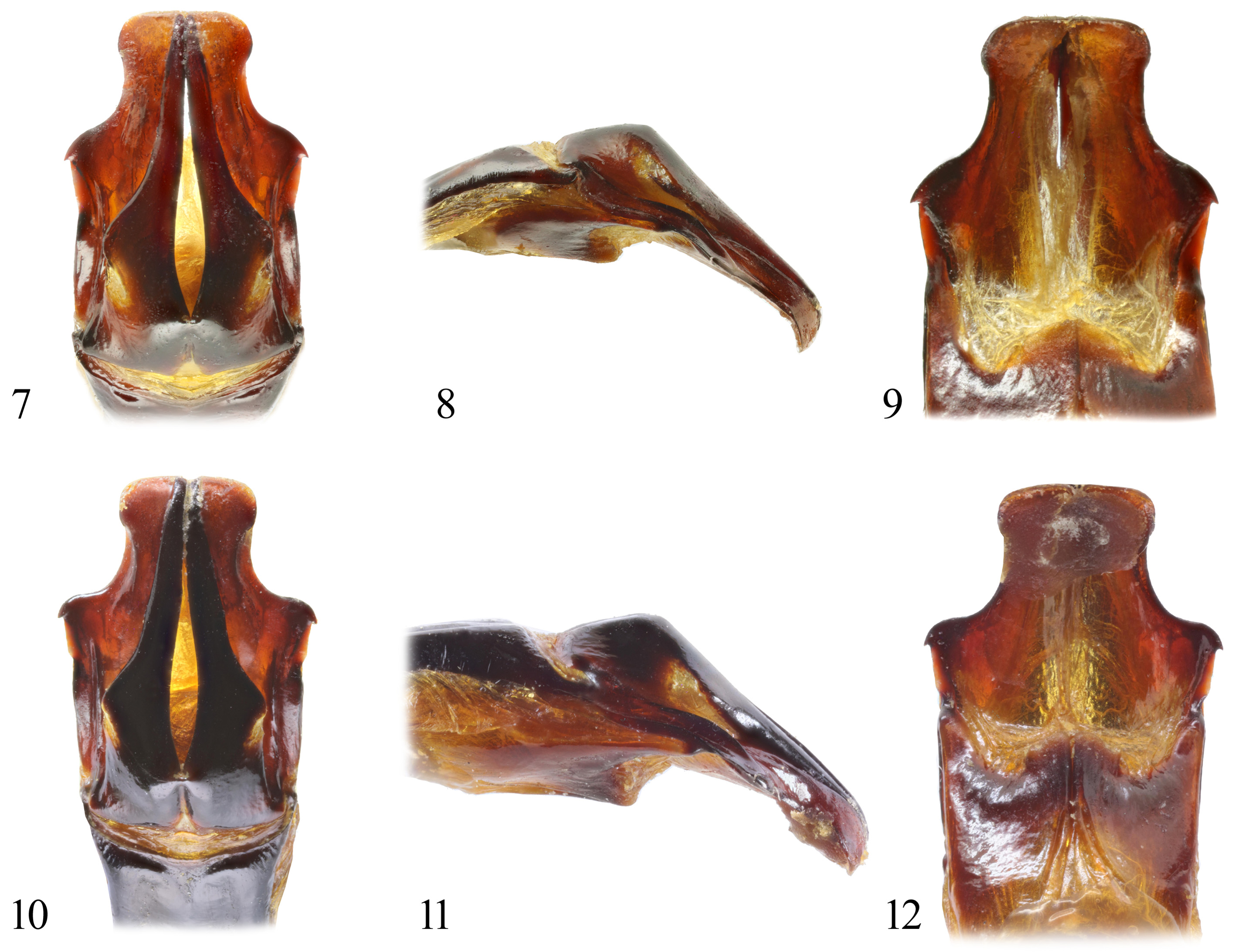
Aedeagus. Parameres ( Figs. 7–9 View FIGURES 7–12 ) slightly shorter than phallobasis, bilobed, sclerotized dorsally and laterally; inner lobe simple, long and narrower apicad; with lateral lobe larger than inner wide and depressed dorsoventrally, with longitudinal lateral depression and oblique lateral carina, with hook-shaped protrusion laterally.
Female ( Figs. 4–6 View FIGURES 1–6 ) differs from male as follows: protibia laterally expanded with all three teeth equally well developed; metatibial spurs slightly more dilated, abdominal ventrites without longitudinal groove.
Measurements. Total body length 24–28 mm, holotype— 26 mm, allotype— 27 mm.
Variability. Type material is only slightly variable in total body length, see measurements above. The color pattern of the dorsal side is relatively stable. Black basal spot on the pronotum has different size and shape. Spots on elytra show different shape and size, often connected together (see Figs. 19, 20 View FIGURES 19, 20 , for detail), mesosternal, metepisternal and metepimeral spots are often absent, lateral spots on abdominal ventrites are variable in shape and size.
Differential diagnosis. Rigout (1989) divided species of the genus Pachnoda into several species groups. Pachnoda iskuulka Král, Sommer & Šípek , new species belongs to the P. abyssinica species group. This group is characterized by a unique aedeagal shape, with parameres ( Figs. 7–18 View FIGURES 7–12 View FIGURES 13–18 ) being always slightly shorter than phallobasis, bilobed, sclerotized dorsally and laterally. The inner lobe is simple, long and narrowed apicad; the lateral lobe is larger than the inner, wide and depressed dorsoventrally, with lateral longitudinal depression and oblique lateral carina, with hook-shaped protrusion laterally. This typical shape of the parameres is also shown in P. iskuulka Král, Sommer & Šípek , new species and in all comparable species (see Figs. 7–18 View FIGURES 7–12 View FIGURES 13–18 and Di Gennaro 2017: pl. 14, fig. 15). The new species is distinctly different from all species so far known from the Horn of Africa (i.e. P. abyssinica abyssinica Reiche, 1847 , P. a. meriteti Di Gennaro, 2017, P. massajae Gestro, 1881 and P. werneri Beinhundner, 1992 ), in a significantly different color pattern of dorsal surface. Only P. iskuulka Král, Sommer & Šípek , new species shows the following combination in the color pattern: head black; pronotum light orange, base with a narrow black strip projecting in variably-shaped spot medially; scutellum black; elytra black with two orange spots just before middle often fused with lateral longitudinal reddish spots, these spots expanded posteriorly inside and often connected to medial reddish spots, praeapical spots reddish and wide ( Figs. 1, 2, 4, 5 View FIGURES 1–6 , 19, 20 View FIGURES 19, 20 , 41, 42 View FIGURES 41–46 ); pygidium with four small white spots along posterolateral margin. The validity of P. iskuulka Král, Sommer & Šípek , new species as a separate species is also supported by the very limited variability in the color pattern of the specimens collected so far (see Figs. 1, 4 View FIGURES 1–6 , 19 View FIGURES 19, 20 for details, altogether 32 specimens from four localities), as well as their offspring (F1) bred during 2018–2019 (see Fig. 20 View FIGURES 19, 20 for details). For this reason, it is almost excluded that these populations would be spontaneous hybrids of two species sensu Rigout (1989: 41 and pl. 11, figs. 5, 6) or Di Gennaro (2017: 60 and pl. 14, figs. 11, 12). Unfortunately, in some of the studies ( Rigout 1989, Di Gennaro 2017) there is not even an indication of where the specimens come from.
Etymology. “ Iskuulka ” means “school” in the Somali language. Most specimens of the new species were caught in the village of Buq near the school where the authors (DK and DS) lived at that time.
Collecting events. Almost all of the type material has been collected from full-grown acacia trees ( Figs. 41, 44 View FIGURES 41–46 ). Beetles appeared on trees at around 11 am and were active even at relatively high temperatures (about 40°C) until dusk. Generally they did not leave the inside of the tree crown, where they were feeding on the flowers and occasionally flying from one flower to another. During such flight bursts, however, they were usually protected by twigs and long thorns. The rest of the time (during the night and the morning when the temperature was lower) they are probably hidden on the ground, under vegetation or in the soil. On all occasions, they were observed together with the abundant cetoniine species Rhabdotis gemella Legrand, 1996 , except during 2017.
Distribution. So far only known from the north-western part of the Cal Madow Mts., at an altitude of approximately 1400–2180 m a.s.l., in the Sanaag region of Somaliland ( Fig. 40 View FIGURE 40 ).
Description of mature (third instar larva) ( Figs. 24–39 View FIGURES 24–33 View FIGURES 34–39 )
Material studied. Six last instar (L3) larvae, reared in laboratory from eggs laid by adults caught in the wild (Buq, 2018—see type material above for details).
Scarabaeiform ( Fig. 24 View FIGURES 24–33 ), maximum length 42–59 mm, cranium yellowish-brow to reddish-brown, glabrous, body whitish. Abdominal segments IX and X fused dorsally, ventrally separated by incomplete groove.
Head capsule ( Figs. 25, 26 View FIGURES 24–33 ). Maximum width 4.0–4.4 mm. Surface of cranium shiny and glabrous, microsculpture linear, sparse and irregular. Cranium yellowish-brown to reddish-brown; antennifer, postclypeus and labrum slightly darker; frontoclypeal border dark brown to black; apices of mandibles black. Cranial chaetotaxy is sum- marized in Table 1 View TABLE 1 . Frontal sutures with a prominent central angle or slightly bisinuate. Epicranial insertions of antennal muscles almost indistinct (visible as only small depressions near middle of frontal suture). Anterior and exterior frontal setae minute. Clypeus trapezoidal to subrectangular, anteclypeal part broad (between 1/3 and 1/2 of entire clypeal area). Postclypeus with one anterior and pair of exterior clypeal setae (one of which is reduced to at least half the size of the other, or even absent). Sclerotization of postclypeus similar to that of frons or epicranium. Stemmata absent.
Antennae ( Fig. 27 View FIGURES 24–33 ). Tetramerous (an I–IV), relative length of antennomeres: an I> an IV ± an II> an III; first antennomere (an I) about length of an II and an III combined. Antennomere III with ventral, apical projection exhibiting single sensory spot. Last antennomere (an IV) usually with two dorsal and three ventral sensory spots and single round, apical sensoric field.
Labrum ( Fig. 25 View FIGURES 24–33 ). Symmetrical, anterior margin trilobed with numerous setae. Clithra present. Dorsal surface in the central part with two prominent setae on each side (one near the center of labrum and one on the lateral margin). Posterior part of labrum with two to six medium sized setae on each side.
Epipharynx ( Fig. 33 View FIGURES 24–33 ). Haptomerum: Zygum convex or slightly angulate and moderately sclerotized with trans- verse, broadly arcuate row of approximately 17–18 stout setae and two rows of another 6–10 stout and prominent setae posterior to the main row. Anterior portion of zygum with several sensilla. Haptomeral process and proplegmata absent. Acroparia: Medial labral lobe about size of lateral lobes or slightly larger. Margin of medial labral lobe with 8–10 setae. Lateral labral lobes with 12–14 setae. Acanthoparia with 6–7 spine-like, slightly recurved setae with enlarged basal areas. Plegmata absent. Chaetoparia asymmetric, right chaetoparia with 91–102 setae in total, left with 66–81 setae. Chaetoparia on each side with a prominent row of 13–15 long and stout setae, extending toward haptolachus; additional one or two rows of similar setae present on right chaetoparia on both sides of the main row. Several more or less irregular rows of slender hair-like setae towards exterior margin of epipharynx. Gymnoparia absent. Dexiotorma, straight, right pternotorma subtle almost absent. Laeotorma distinct, well developed, left pternotorma triangular, large. Haptolachus: Sensorial cone (left nesium) with four pores narrowly conical, sclerotized plate (right nesium) absent. Plate-shaped sclerite consisting of two perpendicular bars situated mediad and to the left of sensorial cone. Area between pedium and anterior portion of haptolachus with approximately 14–16 slender hair-like setae, often continuous with inner parts of left paria; posterolateral part with group of two pore-like setae on each side. Phoba and crepis absent.
Mandibles ( Figs. 28–30 View FIGURES 24–33 ). Asymmetrical, scrobis with 10–13 setae, longitudinal furrow absent. Anterolateral portion of dorsal mandibular surface with two prominent setae and medial pore, another pore found near centre of dorsal mandibular face; 3–10 dorsomolar setae concealed in single rim present on both mandibles. Ventral surface with 3–8 ventromolar setae. Stridulatory area present, with 13–16 transversal ridges in the proximal half of a markedly desclerotised area. Ridges of the stridulatory area uneven, distance between individual ridges about twice in distal ridges compared to proximal ones. Left mandible with four scissorial teeth, apical tooth only slightly falcate and not much larger than others. Second and fourth tooth of left mandible prominent and obtuse. Right mandible with three well developed scissorial teeth. Distal molar lobe of left mandible prominent, much more protruding than remaining parts of mola. Posterior margin of right madibular calyx in medial aspect bilobed, both lobes subtriangular. Dorsal lobe usually about twice as large as ventral. Calyx of left mandible flattened with convex posterior margin. Brustia with 16–21 or 11–17 setae on left and right mandible, respectively.
Maxilla ( Figs. 31, 32 View FIGURES 24–33 , 34 View FIGURES 34–39 ). Dorsal surface of cardo with 5–11 seate, and labacoparia with around 40 setae. Dorsomedial surface of stipes with 23–32 more or less slender hair-like setae and oblique row of 5–7 spine-like stridulatory teeth and anterior truncate process (blunt tubercle). Another 2 or 3 prominent setae located at postero-lateral part of stipes. Ventral surface of stipes and mala distinctly sclerotized, setae prominent and mostly in desclerotised field. Approximately 20 setae present on ventral surface of labacoparia. Galea and lacinia entirely fused forming mala, galeo-lacinial suture indistinct, entirely absent on ventral face. Galear portion of mala with single falcate uncus and 7–8 long and stout hair-like setae in longitudinal rows, and 6–9 slender and shorter setae; lacinia with two unci fused at their base, smaller uncus about half the size of larger one. Maxillary palps tetramerous, penultimate palpomere with two setae.
Hypopharyngeal sclerome ( Fig. 31 View FIGURES 24–33 ). Asymmetrical, truncate process strongly protruding, pointed. Tufts of tegumentary expansions (= phoba, sensu Böving 1936) present on left lateral lobe, at base of truncate process, and proximal to right part of mesolateral margin of hypopharyngeal sclerome, the latter group may be absent in some individuals. Both lateral lobes only feebly sclerotized with approximately 15–20 setae.
Ligula ( Fig. 32 View FIGURES 24–33 ). Dorsal surface with approximately 30 long hair-like setae on each side, a paramedial longitudinal row of 2–3 stout setae near the medial line and a sclerotized oblique row running from central portion of ligula towards apex. Base of ligula with a proximal transverse row of approximately 12 and campaniform setae, interrupted by a paramedial couple of short conical setae. Labial palpi bimerous.
Thorax ( Fig. 24 View FIGURES 24–33 ). Prothorax with single dorsal lobe, meso- and metathorax with three well developed lobes. Dorsi of each thoracic sublobe with approximately 4–6 rows of setae, anterior rows with short setae, setae of posterior row(s) about 3– 5x as long as previous ones. Prothoracic sclerite well sclerotized, covering almost completely lateral portion of prothorax. Prothoracic spiracle ( Fig. 36 View FIGURES 34–39 ) with C-shaped respiratory plate; lobes of respiratory plate well separated with distance between lobes slightly shorter than the maximum diameter of respiratory plate. Respiratory plate with around 35 holes across diameter. All pairs of legs ( Figs. 37–39 View FIGURES 34–39 ) subequal in shape and size. Pretarsi cylindrical with 11–13 setae, claw absent.
Abdomen ( Figs. 24 View FIGURES 24–33 , 35 View FIGURES 34–39 ). Nine-segmented. Dorsum of abdominal segments I–VI with three sublobes, segments VII and VIII with only two. Each sublobe bearing 3–4 rows of setae. Similary to thorax, setae in anterior rows short; posterior row also with medium and long setae. Abdominal spiracles similar to mesothoracic spiracle, all spiracles subequal in size. Dorsum of ultimate abdominal segment (fused segments IX and X) almost evenly covered by sparse medium and long setae, except for the narrow anterior portion without setae. Anal slit transverse emarginated with a dense row of 50–70 setae on ventral anal lip and a similar row of approximately 30 setae on dorsal anal lip.
Raster ( Fig. 35 View FIGURES 34–39 ). Palidium monostichous however, few irregular pali may be scattered around main row. Raster composed of two slightly arched parallel rows of 21–28 pali. Pali flattened distally, with obtusely rounded apices. Septula opened posteriorly, narrow elliptical, about 4 times longer than broad. Tegilla sparsely setose but fused, composed of numerous medium long spiny setae, covering almost completely ventral surface of last abdominal segment.
Note. At this stage it may be expected that we would compare the larval morphology of P. iskuulka Král , Som- mer & Šípek, new species with that of other member of the genus, or with other larvae of the local fauna. However, this is impossible as larvae of only three out of 180 other taxa of Pachnoda are known yet (see Introduction) and they are from completely different regions. Moreover, virtually nothing is known about the immature stages of other Cetoniinae species from the Horn of Africa. Despite this, we are of the opinion that providing information on life cycle and immature stages of this new species adequately complements the description itself, and may be useful in future comparisons.
Remarks on biological cycle in captivity. Seven adults, three males and four females were set into a rearing terrarium at the beginning of September 2018; in the beginning the adults showed minimum activity. However, proper activity started with the installation of a 28W halogen bulb in the breeding enclosure with a 12:12 h light: dark regime. This triggered reproduction and approximately 45 days later larvae of the first and second instar were detected. The adults, meanwhile, continued to feed and be active until the end of November, when the first male specimen eventually died. During the course of their lives their elytral coloration faded considerably. The first L3 was observed in November 2018, only three months after the breeding was established. Pupation was detected during December and at the beginning of January the first adults of the F1 generation emerged, indicating that this species does not undergo any diapause. The larvae fed on the commonly used feeding substrate, but were also observed to feed on debris of ripe fruits and some vegetables (i.e. peelings of cucumber and carrots). Adults of F1 generation preferred cucumber and salad over ripe fruits and beetle jelly.





| NMPC |
National Museum Prague |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Cetoniinae |
|
Genus |