
Ophryotrocha marinae, Zhang & Zhou & Yen & Hiley & Rouse, 2023
|
publication ID |
https://doi.org/ 10.5852/ejt.2023.864.2101 |
|
publication LSID |
lsid:zoobank.org:pub:326759FD-35B4-44C5-B3EF-9E89B6B91106 |
|
DOI |
https://doi.org/10.5281/zenodo.7876068 |
|
persistent identifier |
https://treatment.plazi.org/id/5B66661D-DDF9-425D-860F-412872058C27 |
|
taxon LSID |
lsid:zoobank.org:act:5B66661D-DDF9-425D-860F-412872058C27 |
|
treatment provided by |
Felipe |
|
scientific name |
Ophryotrocha marinae |
| status |
sp. nov. |
Ophryotrocha marinae View in CoL sp. nov.
urn:lsid:zoobank.org:act:5B66661D-DDF9-425D-860F-412872058C27
Ophryotrocha cf. akessoni View in CoL sp. 1 – Goffredi et al. 2017: supplemental, table 1.
Ophryotrocha cf. akessoni View in CoL – Salcedo et al. 2019: 6, table 1.
Etymology
Named in honor of Marina McCowin for her dedication in the study of the fauna associated with seeps and vents. Marina has studied Siboglinidae and Ophryotrocha marinae sp. nov. was notable for being associated in large numbers with the tubes of Oasisia alvinae Jones, 1985 ( Fig. 6A View Fig ).
Material examined
Holotype MEXICO • 1 spec. (fixed in ethanol and the posterior end used for DNA extraction); Gulf of California, Pescadero Basin , active hydrothermal vents; 23.960° N, 108.863° W; depth 3676–3756 m; 18 Apr. 2015; Greg Rouse leg.; collecting event: ROV Doc Ricketts dive 750 (specimens associated with tubes of Oasisia alvinae ); GenBank: OP311750 (COI);, ICML-EMU-13289 , (ex SIO-BIC A14109). GoogleMaps
Paratypes MEXICO • 1 spec. (fixed in ethanol and the posterior end used for DNA extraction); same collection data as for holotype; GenBank: OP311749 (COI), KY701727 (16S), OP311651 (H3); SIO-BIC A6308 GoogleMaps • 1 spec. (fixed in ethanol and the posterior end used for DNA extraction); same collection data as for holotype; GenBank: OP311755 (COI); SIO-BIC A14108 GoogleMaps • 1 spec. (fixed in ethanol and the posterior end used for DNA extraction); same collection data as for holotype; GenBank: OP311751 (COI); SIO-BIC A14110 GoogleMaps • 1 spec. (fixed in ethanol); same collection data as for holotype; SIO-BIC A14111 GoogleMaps • ca 35 specs (10 fixed in formalin, ca 25 fixed in ethanol); same collection data as for holotype; SIO-BIC A14112 GoogleMaps . • 11 specs (5 fixed in paraformaldehyde / glutaraldehyde, 6 fixed in ethanol); Gulf of California, Pescadero Basin , Auka Vent Field, region of P Vent and Z Vent ; 23.96° N, 108.86° W; depth 3648–3671 m; 3 Nov. 2021; Greg Rouse leg.; collecting event: ROV SuBastian dive 475; GenBank: OP561817 (COI); SIO-BIC A14031 . GoogleMaps
Other material
MEXICO • 7 specs (3 fixed in formalin, 4 fixed in ethanol); Gulf of California, Pescadero Basin, Auka Vent Field, Matterhorn area , active hydrothermal vents; 23.95° N, 108.86° W; depth 3650 m; 14 Nov. 2018; Greg Rouse and Ekin Tilic leg.; collecting event: ROV SuBastian dive 193, sample S0193-S4 (suction sampler chamber 4, specimens associated with tubes of Oasisia alvinae ); SIO-BIC A9974 GoogleMaps • 7 specs (2 fixed in formalin, 5 fixed in ethanol); same collection data as for preceding; SIO-BIC A9975 GoogleMaps • 4 specs (fixed in ethanol); same locality as for preceding; 23.95369° N, 108.86231° W; depth 3668 m; 29 Oct. 2021; Greg Rouse leg.; collecting event: ROV SuBastia n dive 470, sample S0470-S3 (suction sampler chamber 3, among tube worms); SIO-BIC A13987 GoogleMaps • 2 specs (1 anterior fragment fixed in ethanol, 1 anterior fixed in formalin and posterior fixed in ethanol and used for DNA extraction); Gulf of California, Pescadero Basin , Auka Vent Field, Z Mound area , active hydrothermal vents; 23.95616° N, 108.86191° W; depth 3688 m; 17 Nov. 2018; Greg Rouse and Ekin Tilic leg.; collecting event: ROV SuBastian dive 196, sample S0196-PC1 (push core 1 at microbial mat with venting); GenBank: OP311752 (COI); SIO-BIC A9987 GoogleMaps • at least 15 specs (3 fixed in formalin, 12 fixed in RNAlater, additional material fixed in ethanol); same locality as for preceding; 23.9561° N, 108.8619° W; depth 3688 m; 21 Nov. 2018; Greg Rouse and Ekin Tilic leg.; collecting event: ROV SuBastian dive 200, samples S0200-S1 and S0200-S2 (suction sampler chambers 1 and 2, microbial mat with venting); SIO-BIC A10029 GoogleMaps • 1 spec. (fixed in ethanol and the posterior end used for DNA extraction); same collection data as for preceding; GenBank: OP311753 (COI); SIO-BIC A14114 • 4 specs (3 fixed in ethanol, 1 anterior fixed in formalin and posterior fixed in ethanol); Gulf of California, Pescadero Basin , Auka Vent Field, oily mat site southeast of Z Vent ; 23.95° N, 108.86° W; depth 3650–3662 m; 5 Nov. 2021; Greg Rouse leg.; collecting event: ROV SuBastian dive 477; SIO-BIC A14045 GoogleMaps • 7 specs (6 fixed in paraformaldehyde, 1 fixed in ethanol); Gulf of California, Pescadero Basin , midway between Auka and JaichMaa ‘ja’ag Vent Fields ; 23.95369° N, 108.86231° W; depth 3663–3687 m; 1 Nov. 2021; Greg Rouse leg.; collecting event: ROV SuBastian dive 473; SIO-BIC A14007 GoogleMaps • 11 specs (2 fixed in formalin, 9 fixed in ethanol); Gulf of California, Pescadero Basin , JaichMaa ‘ja’ag Vent Field, Cavern Tay Ujaa (Big Cave ), active hydrothermal vents; 23.94157° N, 108.85570° W; depth 3675 m; 18 Nov. 2018; Greg Rouse and Ekin Tilic leg.; collecting event: ROV SuBastian dive 197, sample S0197-S6 (suction sampler chamber 6, specimens associated with small tubes of Oasisia alvinae ); SIO-BIC A10002 GoogleMaps • 1 spec. (fixed in ethanol and the posterior end used for DNA extraction); same collection data as for preceding; GenBank: OP311754 (COI); SIO-BIC A14113 • ca 50 specs (fixed in ethanol); Gulf of California, Guaymas Basin , sedimented hydrothermal vents; 27.016° N, 111.410° W; depth 2012 m; 13 Nov. 2009; Anna-Louise Reysenbach leg.; collecting event: HOV Alvin dive 4558; SIO-BIC A14115 GoogleMaps • 1 spec. (fixed in ethanol and the posterior end used for DNA extraction); same collection data as for preceding; GenBank: OP311756 (COI); SIO-BIC A14116 • 1 spec. (fixed in ethanol and the posterior end used for DNA extraction); same collection data as for preceding; GenBank: OP311757 (COI); SIO-BIC A14117 • 1 spec. (fixed in ethanol and the posterior end used for DNA extraction); same collection data as for preceding; SIO-BIC A14118 • ca 15 specs (fixed in ethanol); Gulf of California, Guaymas Basin , sedimented hydrothermal vents; 27.015° N, 111.410° W; depth 2010 m; 15 Nov. 2009; Anna-Louise Reysenbach leg.; collecting event: HOV Alvin dive 4560; SIO-BIC A14119 GoogleMaps • 1 speci. (fixed in ethanol and the posterior end used for DNA extraction); same collection data as for preceding; GenBank: OP311758 (COI); SIO-BIC A14120 • 1 spec. (fixed in ethanol and the posterior end used for DNA extraction); same collection data as for preceding; GenBank: OP311759 (COI); SIO-BIC A14121 • 1 spec. (fixed in ethanol and the posterior end used for DNA extraction); same collection data as for preceding; GenBank: OP311760 (COI); SIO-BIC A14122 .
Description
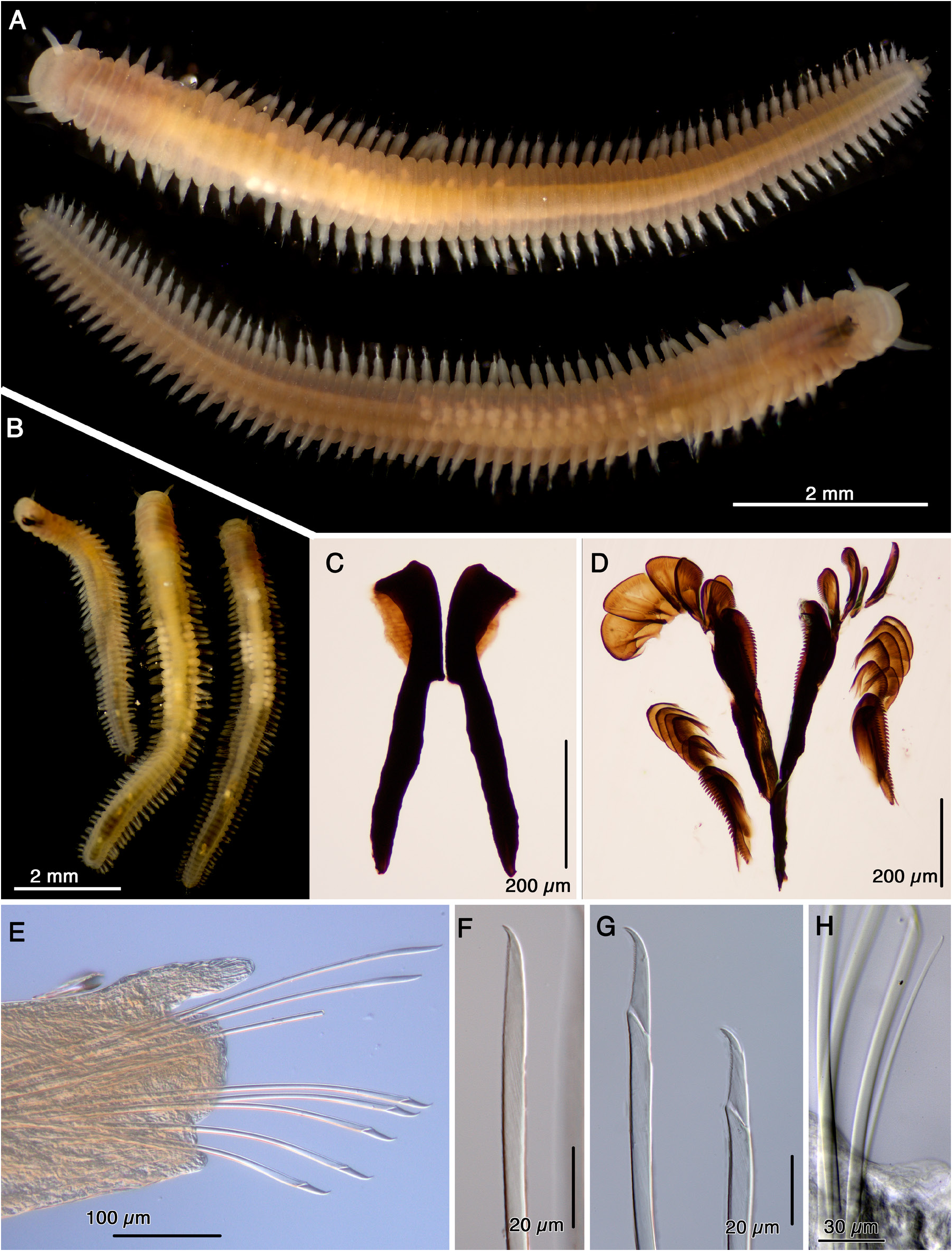
In life, golden color with white eggs mid-body ( Fig. 7A–B View Fig ), opaque white after preservation. Body length 10.5 mm, with 50+ segments, slightly dorso-ventrally compressed, widest anteriorly, gradually tapering posteriorly ( Fig. 7A–B View Fig ). Prostomium rounded, wider than long, posterior medial area slightly raised, with paired digitiform antennae, tapering distally, inserted dorsally, paired digitiform palps similar in length with antennae, inserted ventral-laterally ( Fig. 7A View Fig ). Peristomium with two rings, subequal in length to the following segments ( Fig. 7A View Fig ). Eyes not visible. Mandibles heavily sclerotized, with rod-like shafts, sub-triangular shape cutting plates, with single blunt peak anteriorly, lateral wings weakly sclerotized ( Fig. 7C View Fig ). Maxillae P-type, forceps comb-like, with large main fang. Four rows of free denticles, the posterior-most free denticles (D1) comb-like, like the forceps, other free denticles shovel-shaped with fine teeth, D2–D4 smaller than D5–D7 ( Fig. 7D View Fig ). Parapodia uniramous, acicular lobe with rounded distal margin, dorsal cirri long digitiform, extending beyond distal margin of acicular lobe, ventral cirri short and stubby ( Fig. 7E View Fig ). Supra-acicular chaetae simple, distally serrated, tapering into a main fang ( Fig. 7F View Fig ). Sub-acicular chaetae compound, with serrated blades ( Fig. 7G View Fig ). Sub-acicular chaetal lobe usually with one simple chaeta ( Fig. 7H View Fig ). Pygidium with two digitiform cirri inserted laterally ( Fig. 7A View Fig ).
Distribution
Known from Gulf of California hydrothermal vents of the Pescadero Basin at over 3500 m and the Guaymas Basin sedimented vents at ~ 2000 m. Found in huge numbers on tubes of Oasisia ( Fig. 6A View Fig ), or on microbial mats near active flow ( Fig. 6B View Fig ).
Remarks
This species was initially reported in Goffredi et al. (2017) as Ophryotrocha cf. akessoni sp. 1 with a partial DNA sequence for mitochondrial 16S rRNA lodged on GenBank (KY701727). Ophryotrocha marinae sp. nov. most closely resembles Ophryotrocha jiaolongi described from hydrothermal vents of the Indian Ocean, including sharing distinctive mandibles that distinguish these two taxa from all other Ophryotrocha . Morphologically, O. marinae differs from O. jiaolongi based on body color, the form of antennae and palps, and in lacking a median pygidial papilla ( Table 3 View Table 3 ). In life, O. marinae is golden while O. jiaolongi is white and translucent. Ophryotrocha marinae also has antennae and palps that are distally tapering and longer than the length of prostomium, while in O. jiaolongi antennae and palps are shorter than the length of prostomium. The minimum COI uncorrected distance obtained between O. marinae and O. jiaolongi specimens was relatively small at 3.73% ( Table 2 View Table 2 ). Other relatively small distances that are currently known are up to 4.8% between O. notoglandulata Pfannenstiel, 1972 and O. japonicus Paxton and Åkesson, 2010 (both Japanese taxa) and 6.1% between O. flabella and O. globopalpata (both from deep waters of the eastern Pacific). Given the morphological differences between O. marinae and O. jiaolongi , the reciprocal monophyly based on numerous COI sequences and vast geographic separation, we regard them here as separate species. One notable difference between O. jiaolongi and O. marinae was that the latter species showed four rows of maxillae compared to the two rows in O. jiaolongi found by Zhang et al. (2017). As discussed above for O. charlottae sp. nov. it is possible that the outermost pair of rows represent molted jaws as has been observed in other Ophryotrocha by Paxton (2004).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ophryotrocha marinae
| Zhang, Dongsheng, Zhou, Yadong, Yen, Nicole, Hiley, Avery S. & Rouse, Greg W. 2023 |
Ophryotrocha cf. akessoni
| Salcedo D. L. & Soto L. A. & Paduan J. B. 2019: 6 |