
Myzophyllobothrium Shipley and Hornell, 1906
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4999.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:051E68AE-6A5B-44AC-8F90-AEECB8997883 |
|
persistent identifier |
https://treatment.plazi.org/id/2A0987A5-FFCD-FFF2-FF7A-3A6F2070F83A |
|
treatment provided by |
Plazi |
|
scientific name |
Myzophyllobothrium Shipley and Hornell, 1906 |
| status |
|
Myzophyllobothrium Shipley and Hornell, 1906
(Syns. Mixophyllobothrium Shinde and Chincholikar, 1980 ; Myzocephalus Shipley and Hornell, 1906 ; Rhoptrobothrium Shipley and Hornell, 1906 )
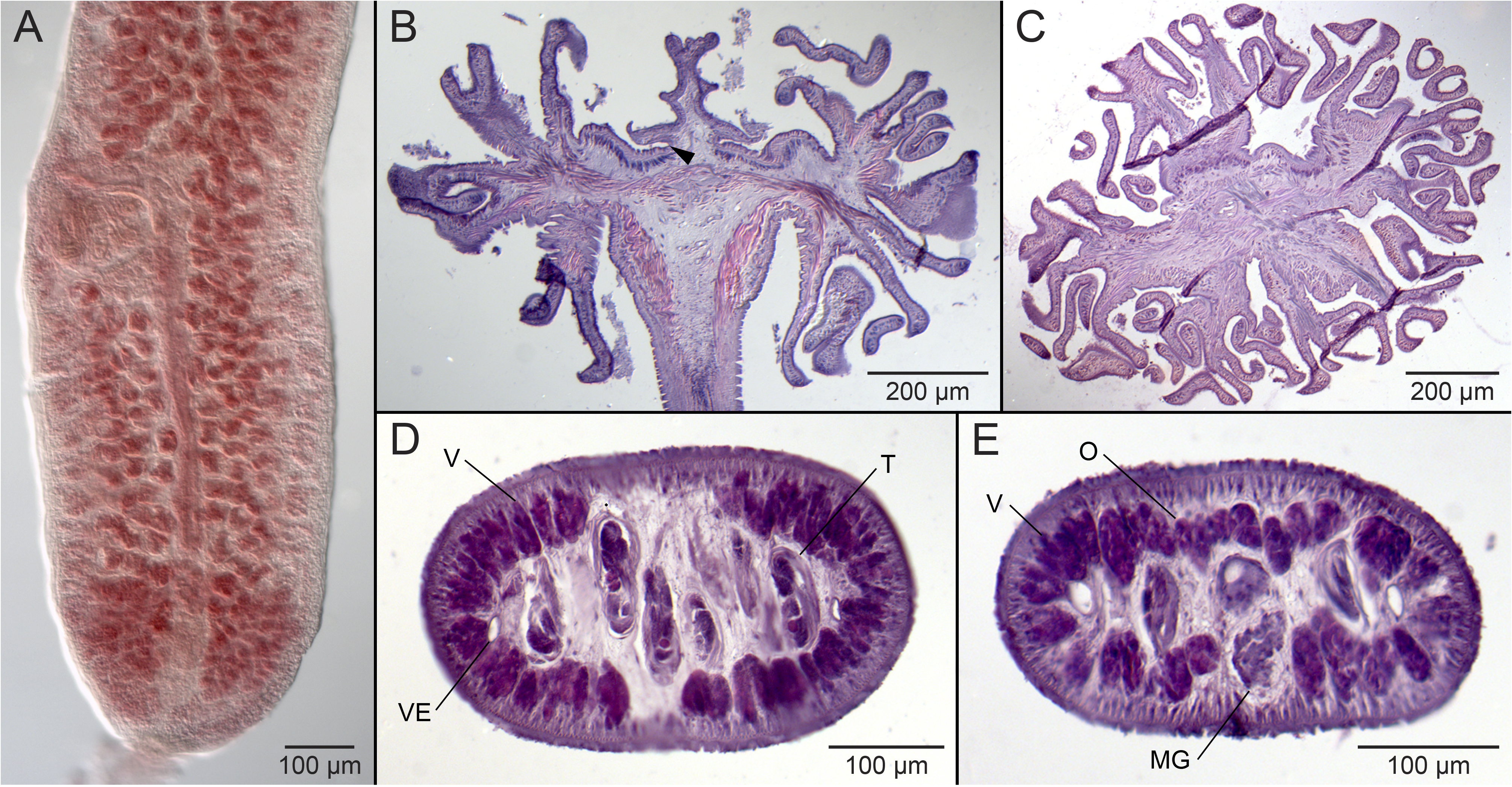
Generic diagnosis. Rhoptrobothriidae . Worms euapolytic. Scolex with four acetabula and cephalic peduncle. Acetabulum bothridiate in form, with two loculi; anterior loculus undivided, with or without postero-lateral extensions; posterior loculus undivided. Cephalic peduncle bearing four stalked remi; remus with terminal primary areola, with or without subterminal secondary areola. Neck absent. Proglottids craspedote or acraspedote. Testes numerous, anterior to ovary or ovarian bridge, arranged essentially in single dorso-ventral layer. Vas deferens coiled, medial to cirrus sac. Cirrus sac pyriform, containing coiled cirrus. Cirrus armed with spinitriches. Ovary H-shaped in frontal view, tetralobed in cross section. Vagina weakly sinuous, opening into genital atrium anterior to cirrus sac; vaginal sphincter absent. Genital pores lateral, irregularly alternating. Uterus medial, saccate, extending to cirrus sac. Vitellarium follicular; follicles arranged in two lateral bands; each band consisting of one to many columns of follicles, extending entire length of proglottid, may be interrupted by ovary. Parasites of Aetobatidae and Myliobatidae . Indian and western Pacific Oceans.
Type species: Myzophyllobothrium rubrum Shipley and Hornell, 1906 .
Additional species: Myzophyllobothrium chongi ( Jensen and Caira, 2006) n. comb., Myzophyllobothrium gambangi ( Jensen and Caira, 2006) n. comb., Myzophyllobothrium limae ( Jensen and Caira, 2006) n. comb., Myzophyllobothrium myliobatidis ( Shipley and Hornell, 1906) Southwell, 1925 , Myzophyllobothrium nagasawai n. sp., and Myzophyllobothrium narinari ( Shipley and Hornell, 1906) n. comb.
Species inquirendum: Myzophyllobothrium okamuri ( Shinde and Chincholikar, 1980) n. comb.
Remarks. The synonymization of Mixophyllobothrium , Myzocephalus , and Rhoptrobothrium with Myzophyllobothrium is supported by both morphological and molecular criteria. With respect to morphology, although the original descriptions suggest that the scolices of these genera differ dramatically from one another, examination of material of M. narinari and M. nagasawai n. sp., the latter of which most closely resembles M. rubrum , with light and scanning electron microscopy reveals fundamental commonalities across these taxa. All bear a scolex that consists of four bothridia each of which bears an anterior and a posterior loculus, giving the bothridia a biloculate appearance. Although Myzophyllobothrium was described as possessing “four almost terminal suckers” ( Shipley and Hornell 1906; pg. 48), the biloculate appearance of the bothridia was illustrated in their fig. 18 and was confirmed by Joyeux and Baer (1961; fig. 290) after examination of a co-type specimen of M. rubrum . The remaining differences are in the four remi that extend from the cephalic peduncle posterior to the bothridia. However, these differences can be considered to be essentially just variations in the width and extent of folding of the remi. In M. narinari and M. rubrum the remi are either much wider than, or nearly as wide as, they are long; in all four species formerly assigned to Rhoptrobothrium , the remi are longer than wide. The remi of M. narinari are highly folded; those of M. rubrum and M. nagasawai n. sp. are less folded; in all four species formerly assigned to Rhoptrobothrium , the remi are weakly or essentially unfolded. These taxa also share a number of notable similarities in proglottid anatomy. In all seven valid species, the cirrus sac is relatively large, extending to the midline of the proglottid, and is located near the middle of the proglottid. All species also possess a post-poral field of testes and a uterus that extends anteriorly in mature proglottids only to the level of the cirrus sac. In all species, the ovary is lobulated, H-shaped in frontal view, and tetralobed in cross section. The main differences among species are in testes number, number of columns of vitelline follicles, and whether the columns are interrupted by the ovary. The microthrix patterns on the scolices of the six species that have been examined with SEM are also very consistent. Little variation exists across surfaces of the scolex; all surfaces of the scolex of all species bear small, slender gladiate spinitriches and capilliform filitriches, albeit of varying relative densities.
The terminology we have applied here to the configuration of the bothridial portion of the scolex differs from that used by Jensen and Caira (2006) for species of Rhoptrobothrium . These authors followed the terminology of Caira et al. (1999; 2001) and described the bothridia (e.g., Jensen and Caira, 2006, pg. 191) as bearing a “specialized anterior region in form of loculus and undivided main loculus”. This terminology was employed to reflect the homology of the different regions of the bothridia of these species with those of other genera. For clarity, we have simplified this terminology here and refer to the bothridia as bearing an anterior and a posterior loculus (i.e., biloculate bothridia). The homologies described by Jensen and Caira (2006) stand; but this revised terminology simplifies comparisons with previous descriptions of members of this group.
Among the 21 valid genera of phyllobothriideans (see Ruhnke et al. 2017; Caira et al. 2021), Myzophyllobothrium most closely resembles Ruhnkebothrium Caira, Bueno, and Jensen, 2021 and Thysanocephalum . All three genera appear to exhibit a scolex that is divided into two regions—a narrow anterior region bearing four biloculate bothridia and an extensive, often convoluted, posterior region, that also is divided into four parts (see Linton 1892, Caira et al. 2021). These similarities are, however, superficial. In Thysanocephalum and Ruhnkebothrium , the scolex has been determined to consist of four bothridia each of which bears a single loculus at its narrow anterior end and an extensive, convoluted posterior loculus ( Caira et al. 1999, 2021, respectively). In contrast, the four biloculate bothridia of Myzophyllobothrium are completely independent structures from the remi, which are extensions of the cephalic peduncle (see Jensen and Caira, 2006 and Fig. 4C View FIGURE 4 ). The independence of these three genera from one another was also supported by the results of the molecular phylogenetic analysis of Caira et al. (2014) in which the representatives of Thysanocephalum , Ruhnkebothrium (referred to as New genus 10), and Myzophyllobothrium (referred to as Rhoptrobothrium and Myzocephalus ) grouped with other taxa in different parts of the tree.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.