
Myrmeleotettix pallidus (Brunner von Wattenwyl, 1882)
|
publication ID |
https://doi.org/10.11646/zootaxa.4895.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:002F9E9D-43AA-4CD3-89FB-FD41EEEE4B18 |
|
DOI |
https://doi.org/10.5281/zenodo.4386344 |
|
persistent identifier |
https://treatment.plazi.org/id/03D81D4E-FFC9-0E12-FF4E-FE0A4A321840 |
|
treatment provided by |
Plazi |
|
scientific name |
Myrmeleotettix pallidus (Brunner von Wattenwyl, 1882) |
| status |
|
Myrmeleotettix pallidus (Brunner von Wattenwyl, 1882)
Distribution. South-eastern part of European Russia, Kazakhstan, Altai, south-western Siberia.
Material. Kazakhstan: 4. Akmola region , ab. 40 km NW of Ereimentau, steppe near Baysary , 51°59.3’ N, 72°42.1’ E, 03.07.2019, song recording in 1 ³; GoogleMaps 5. Pavlodar region, ab. 48 km W of Ekibastuz, steppe ab. 3 km W of Schiderty, 51°42.9’ N, 74°38.5’ E, 04.07.2019, song recording in 1 ³; GoogleMaps 9. Pavlodar region, environs of Irtyshsk, 53°20.2’ N, 75°25.8’ E, 05.07.2019, song recordings in 3 ³ GoogleMaps .
References to song. Savitsky, 2005, 2009: recordings from Russia ( Volgograd region), calling and courtship songs.
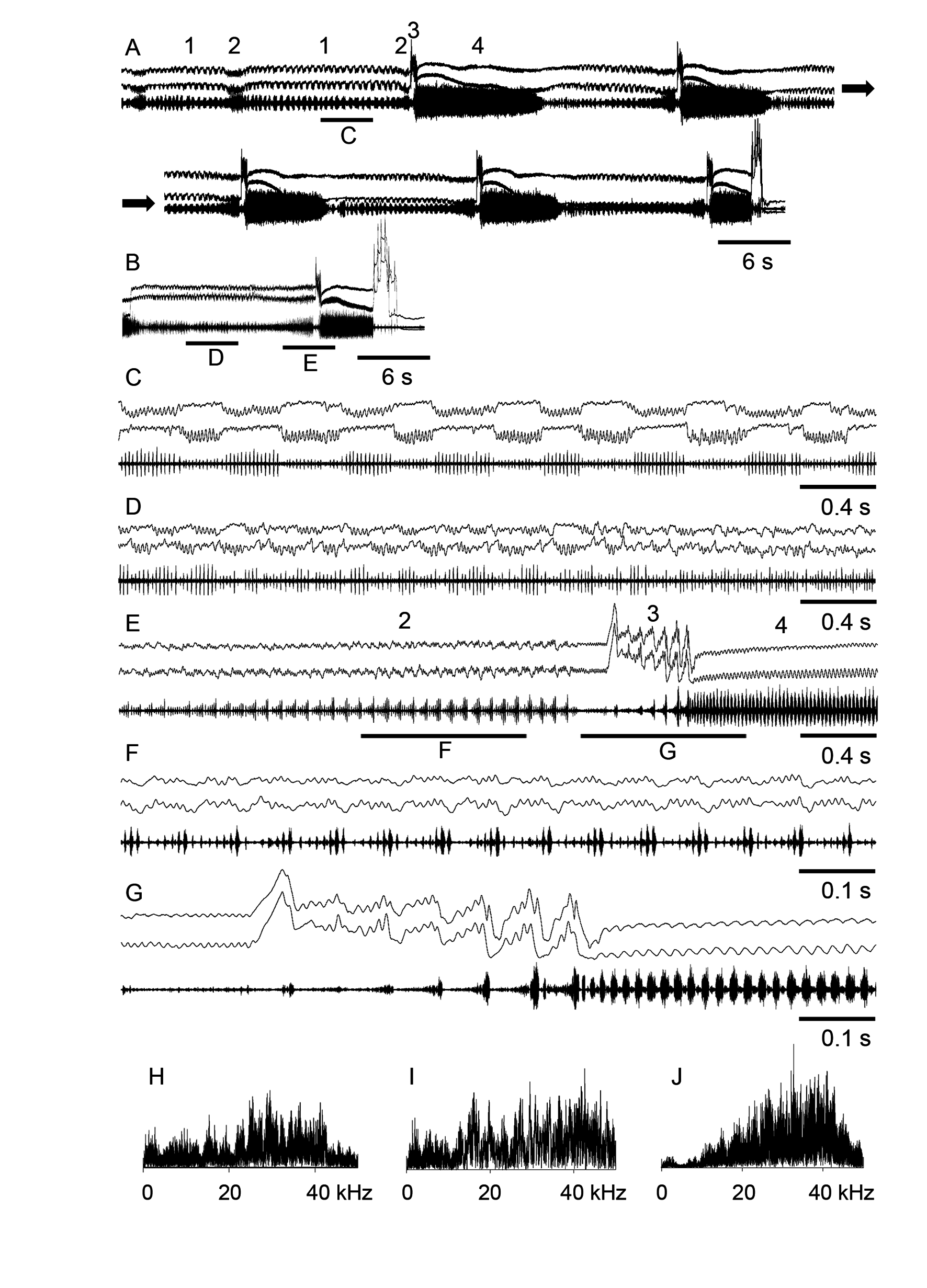
Song. The courtship song starts with alternating movements of the two hind legs (element 1, Fig. 5 A, C View FIGURE 5 ). One leg produces from 10 to 15 low-amplitude up and down movements, whereas the other leg doesn’t move. Next, the legs change the roles. Despite the two legs demonstrate similar movement patterns, one leg produces the pulses of larger amplitude than the other leg. Thus, the syllables of larger pulses alternate with the syllables of the smaller pulses. Sometimes the legs do not show regular alternation of vibrations and pauses, in which case the pulse amplitude varies randomly ( Fig. 5 D View FIGURE 5 ). The pulse repetition rate is typically about 40–55/s. This element lasting 6–10 s follows by the element 2 of the courtship song, which is similar to the calling song ( Fig. 5 E View FIGURE 5 ). It lasts 1–3 s and consists of the syllables repeated at the rate of about 10–20/s. Each syllable is usually composed of 7–10 pulses that reach maximum intensity in the end of each syllable. During the element 2, the legs are moved almost synchronously ( Fig. 5 F View FIGURE 5 ). The two elements alternate during several minutes. Thereupon, the element 2 is followed by the element 3 that is accompanied by a visual display ( Fig. 5 E, G View FIGURE 5 ). The legs are lifted to an extra high position and are moved according to a specific pattern, which is nearly similar to the pattern of the element 2, but the very movements are larger in amplitude. The rate of the principal leg-movements is about 16/s, and superimposed on these movements there are the small-amplitude vibrations of the rate of about 90–110/s. The amplitude of syllables gradually increases so that the last syllable is the loudest. During the element 3, a male is also conspicuously moving his clubbed antennae in horizontal plane ( Fig. 6 View FIGURE 6 ). Immediately after the element 3, the element 4 is emitted. It is produced by synchronous, low-amplitude movements of the legs and lasts about 6–10 s. A low-amplitude pulse is produced during up-stroke, whereas a high-amplitude pulse is generated during down-stroke. Sometimes, the low-amplitude pulses are almost invisible on the oscillogram. The syllable rate in the element 4 is about 50–63/s. After the element 4, there is a sequence of the elements 1–4; several sequences are followed by precopulatory movements and an attempt to copulate with a female ( Fig. 5 A, B View FIGURE 5 ). The frequency spectra of the sound produced during various parts of the courtship song are different. The frequency spectrum the element 2 has the main maxima around 25 and 28 kHz; the spectrum of the element 3 has many periodical peaks in the broad range of 15–45 kHz; the spectrum of the element 4 has main components between 25 and 45 kHz ( Fig. 5 View FIGURE 5 H–J).
Comparative remarks. Our recordings of the courtship song in M. pallidus from Kazakhstan do not basically differ from the recordings of this species from Volgograd region ( Savitsky, 2005). However, there are some minor differences. First, we found the difference in the fine temporal structure of the syllables in the song element 4. In the song recordings from Volgograd, the pulses of the element 4 are of similar amplitude, whereas the analysis of the recordings from Kazakhstan reveals the alternation of the low-and the high-amplitude pulses. In addition, the syllables of the element 3 are the loudest ones in the recordings from Volgograd ( Savitsky, 2005), whereas in our recordings, the element 4 is the loudest. This can be explained by different frequency characteristics of these two elements. A more broad-band sound is typical for the syllables of the element 3, whereas dominant frequencies of the element 4 shift to the ultrasound band ( Fig. 5 I, J View FIGURE 5 ). The recordings of Savitsky (2005) were made in the field with cassette recorders; the upper frequency limit was not higher than 15 kHz. This could explain why the amplitude ratio of the elements 3 and 4 of the courtship song might be distorted in the recordings from Volgograd. The distortion of the frequency characteristics may also explain the difference in the pulse structure of the element 4 between the recordings from Volgograd region and Kazakhstan. Finally, Savitsky (2005) does not mention any movements with antennae during courtship.
The movements with antennae during courtship are typical for all three species of Myrmeleotettix studied so far. In M. maculatus (Thunberg, 1815) and M. antennatus (Fieber, 1853) , antennae are moved in longitudinal plane—back and forth over the head ( Bull, 1979; Berger & Gottsberger, 2010; Vedenina & Shestakov, 2014). In some other grasshopper species of the genera Stenobothrus , Gomphocerus and Gomphocerippus ( Elsner, 1974; Ostrowski et al., 2009) antennae are also swung mainly in longitudinal plane. Strokes with antennae are supposed to originate from the disturbance reactions ( Willey & Willey, 1969; Berger & Gottsberger, 2010), which can be frequently observed when animals are disturbed or touched by other animals or an experimenter. In this case the animals raise antennae into an opposite side of the disturbing interference, which usually means backwards. By contrast, in M. pallidus antennae are moved in horizontal plane (from side to side). Such pattern of the antennae movements seems to be unique among Gomphocerinae .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Gomphocerinae |
|
Genus |
Myrmeleotettix pallidus (Brunner von Wattenwyl, 1882)
| Vedenina, Varvara, Sevastianov, Nikita & Tarasova, Tatiana 2020 |
Gomphocerippus
| Roberts 1941 |
Myrmeleotettix
| Bolivar 1914 |
Stenobothrus
| Fischer 1853 |
Gomphocerinae
| Fieber 1853 |