
Mycale (Mycale) lobos, Turner & Lonhart, 2023
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5318.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:88714F9C-0EE5-4295-9988-3CEEF242489D |
|
DOI |
https://doi.org/10.5281/zenodo.8162417 |
|
persistent identifier |
https://treatment.plazi.org/id/464C8784-426B-FFC2-FF1D-FE12FB03FEC1 |
|
treatment provided by |
Plazi |
|
scientific name |
Mycale (Mycale) lobos |
| status |
sp. nov. |
Mycale (Mycale) lobos View in CoL sp. nov.
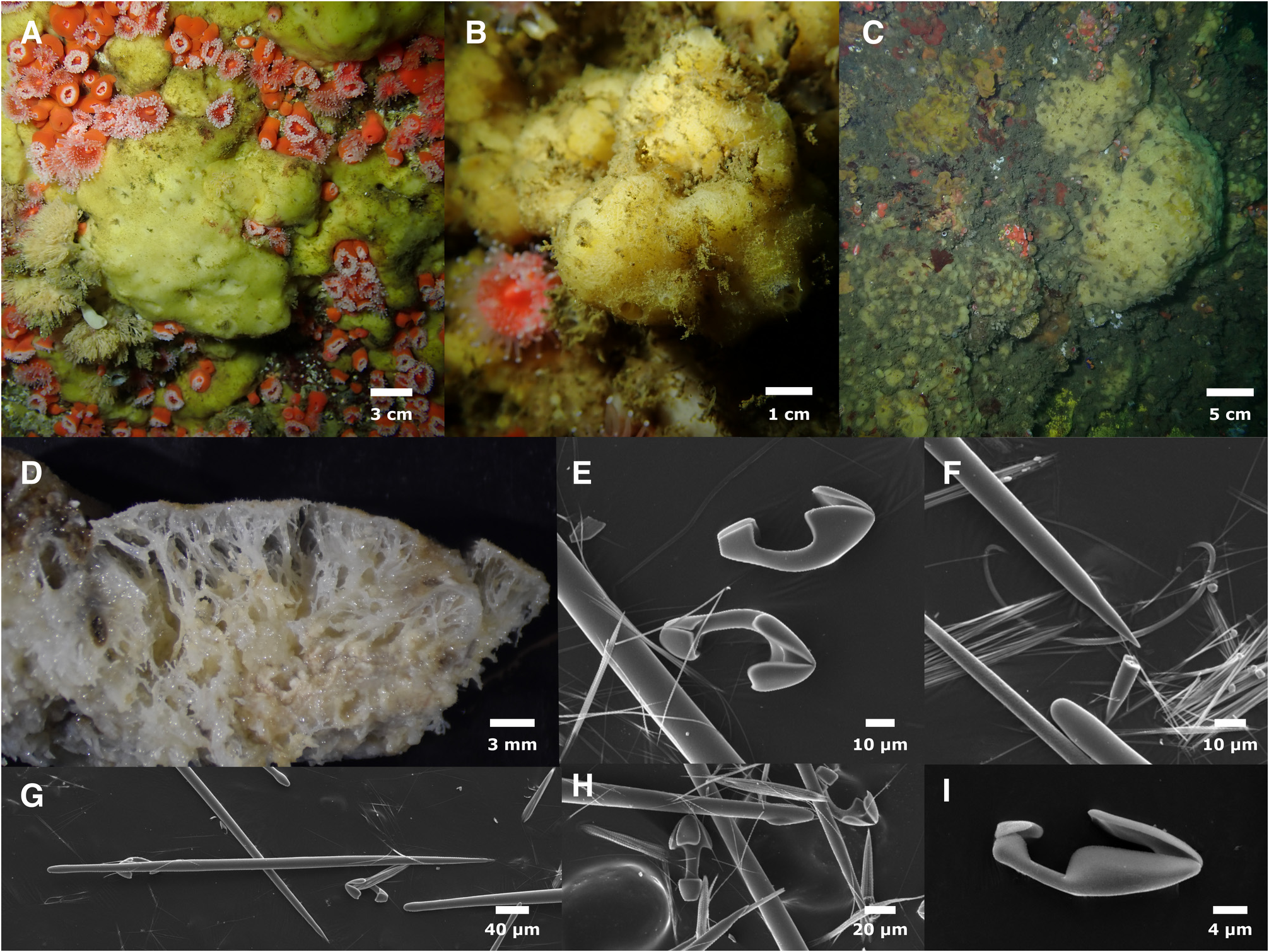
Figure 13 View FIGURE 13 .
Material examined. Holotype: CASIZ236659 / IZC00048460 , Inner Carmel Pinnacle, (36.55852, -121.96820), 10– 24 m, 9/22/21; GoogleMaps paratypes: SBMNH700909 About SBMNH , Point Pinos, Pacific Grove, (36.64183, -121.93060), 9–18 m, 8/9/21; GoogleMaps SBMNH700920 About SBMNH , Whaler's Cove, Point Lobos, Carmel, (36.52172, -121.93894), 6–15 m, 11/23/19; GoogleMaps IZC00048458 , Dino Head, Point Loma, San Diego, (32.68808, -117.27080), 21–26 m, 9/19/20; GoogleMaps IZC00048459 , Nudi Wall, Point Loma, San Diego, (32.69872, -117.27580), 18–28 m, 5/16/21, GoogleMaps SBMNH700923 About SBMNH , Otter Cove, Pacific Grove (36.63110, -121.91840), 9–15 m, 8/9/21. GoogleMaps
Etymology. Name inspired by Point Lobos, where the first sample was found.
Morphology. Thickly encrusting or massive; samples are 2–3 cm thick, but some sponges were up to at least 10 cm thick in life. Irregular in thickness, often appearing as an irregular, featureless blob without visible surface patterning or oscules. Oscules sparse in some samples but more common in others, approximately 2–4 mm in diameter, sometimes found in small clusters. Surface slightly hispid, often with ensnared silt and other debris; lacks surface furrows known to characterize M. lingua ( Bowerbank, 1866) and M. toporoki Koltun, 1958 . Ectosome usually pale yellow but sometimes white in life. Large subectosomal vacuoles; in several samples, this space was packed with yellow spheres that appeared to be developing larvae, up to 400 μm in diameter. All tissues white when preserved.
Skeleton. Reticulate, anastomosing spongin fibers, cored with styles, rise to the surface of the sponge and support an unstructured mat of styles in the ectosomal skeleton. Large anisochelae occur in rosettes. Trichodragmas are extremely abundant.
Spicules. Mycalostyles, anisochelae (2 sizes), sigmas (2 sizes), trichodragmas.
Mycalostyles: Thickest in center of shaft, but usually subtylote due to constriction ("neck") below head. Heads generally oblong and sometimes absent; rarely transformed into oxeas. In one sample ( IZC00048459 ) many are polytylote; this was seen rarely in other samples. Significantly shorter in ectosome than choanosome (Wilcoxon rank–sum test p=7.18e–10). Holotype ectosome: 381–471–671 x 7–14–20 μm (n=103). Holotype choanosome: 377–542–631 x 6–13–18 μm (n=76). If spicule preparations are done from combined ectosomal and choanosomal tissue, length distribution is bimodal, with peaks at ~450 and ~550 μm. All samples and tissues pooled: 342–496– 671 x 6–13–21 μm (n=401).
Large anisochelae: shaft distinctly curved in profile. Holotype 44–54–63 μm (n=31). All samples pooled 31– 51–63 μm (n=158). Shaft length to alae length ratio averages 0.35.
Small anisochelae: Holotype 13–17–24 μm (n=22), all samples pooled 9–17–24 μm (n=169).
Small sigmas: uncommon. All samples pooled 15–18–23 μm (n=5).
Large sigmas: Holotype 45–61–72 μm (n=15), all samples pooled 43–57–72 μm (n=38).
Trichodragmas: abundant. Holotype 33–62–79 x 6–10–13 μm (n=19), all samples pooled 31–58–76 x 4–9–14 μm (n=28), individual raphides 1–2 μm in width.
Distribution and habitat. This species is common but patchily–distributed on subtidal rocky reefs around the Monterey Peninsula (Central California), where it was seen at 5 of 13 reefs investigated. It was also abundant at two sites around the Point Loma kelp forests (San Diego, Southern California), but not seen at the other 6 reefs investigated in that area. Both Point Loma sites were among the deepest sites investigated, with maximum depths of 26– 28 m. We therefore hypothesize that it is common in Central California on shallow subtidal reefs, and also common in Southern California, but at slightly deeper reefs.
Remarks. The ectosomal skeleton and the lack of modified chelae serve to place this species in the subgenus Mycale (Mycale) . Though there are many species of Mycale known from the Eastern Pacific, the curved shaft of the large anisochelae differentiate this species from all but two ( Carballo & Cruz-Barraza 2010). Mycale darwini Hajdu & Desqueyroux-Faúndez 1994 , known from the Galapagos Islands, has curved anisochelae, but grows as a thin crust that is a poor match to the current species. Mycale toporoki Koltun, 1958 is more similar. This species was described from Northern Russia, and may also occur in the Aleutian Islands, Pacific Northwest, and possibly California ( Lee et al. 2007; Lehnert et al. 2006). A similar sponge from the North Atlantic, M. lingua ( Bowerbank, 1866) , has also been reported in the North Pacific by some authors, though others have suspected these reports actually refer to M. toporoki ( Bakus 1966; Lee et al. 2007; Lehnert et al. 2006). Both M. lingua and M. toporoki are characterized by surface grooves that are "obvious in living specimens" ( van Soest & Hajdu 2002). All sponges of the new species were examined alive and nothing resembling these grooves were seen in any of the specimens. Moreover, the sizes of the styles and both classes of chelae of the new species are much smaller than M. toporoki and M. lingua . Reports for both of these named species have the smaller size class of chelae as 20–30 μm and the larger size class as 60–100 μm. Mycale lobos sp. nov., in contrast, has chelae 11–24 μm and 31–63 μm. Despite measuring 327 chelae, and extensively searching for larger spicules, no chelae longer than 63 μm could be found. No styles longer than 622 μm could be found in the new species either, while the two previously described species have styles up to 800 μm or over 1000 μm, respectively.
This species is not shown in the phylogenies, as we failed to generate genetic data for this species or the congeneric M. psila below; this is perhaps due to primer mismatches.
Many specimens of M. lobos sp. nov. can be tentatively identified in the field based on gross morphology, albeit mostly by the lack of features. Sponges growing as large, featureless blobs, pale yellow in color, with hispidity collecting debris have thus far proved to be this species upon examination. Tearing the sponge in the field, to look for large cavities filled with developing larvae, would increase confidence of field identification.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |