
Milnesium cassandrae, Moreno & Roszkowska & García & Flores & Kaczmarek, 2019
|
publication ID |
https://doi.org/10.11646/zootaxa.4691.5.5 |
|
publication LSID |
lsid:zoobank.org:pub:0684AE18-0510-4F7B-B75D-AE5177FBF2A2 |
|
persistent identifier |
https://treatment.plazi.org/id/0684AE18-0510-4F7B-B75D-AE5177FBF2A2 |
|
taxon LSID |
lsid:zoobank.org:act:0684AE18-0510-4F7B-B75D-AE5177FBF2A2 |
|
treatment provided by |
Plazi |
|
scientific name |
Milnesium cassandrae |
| status |
sp. nov. |
Milnesium cassandrae sp. nov.
( Table 2 View TABLE 2 , Table 3 View TABLE 3 , Figure 1 View FIGURE 1 a–e, Figure 2 View FIGURE 2 a–e)
Type material: Holotype (female) and 47 paratypes ( 32 adult females, 13 hatchlings, two juveniles of indeterminate sex), 25 eggs and nine exuviae from piedmont of Sierra Las Mitras ( 25°44′55.57′′N, 100°24′11.19′′W, ca. 632 m asl), Monterrey, Nuevo León, Mexico. Lichen ( Physcia stellaris ) on trees ( Parkinsonia aculeta and Celtis pallida ), coll. Antonio Moreno - Talamantes, 7 August 2014. GoogleMaps
Additional material: Thirteen females from a lichen sample and five eggs in exuviae from San Agustin Park ( 25°39′18.63′′N, 100°20′34.51′′W, ca. 605 m asl), San Pedro Garza García Municipality, Nuevo Leon. Mexico. Lichen ( Nephroma sp.) on trees ( Fraxinus sp. and Quercus sp.), coll. Antonio Moreno - Talamantes, 6 November 2013. Four females from a piedmont of Sierra Las Mitras ( 25°44′45.98′′N, 100°25′27.68′′W, ca. 700 m asl), Monterrey, Nuevo Leon, Mexico. Lichen ( Physcia stellaris ) on tree ( Quercus virginiana ), coll. Antonio Moreno - Talamantes, 8 November 2014. Six females from piedmont of Sierra Las Mitras ( 25°44′38.55′′N, 100°25′31.65′′W, ca. 719 m asl), Monterrey, Nuevo Leon, Mexico. Lichen ( Physcia stellaris ) on tree ( Quercus virginiana ), coll. Antonio Moreno - Talamantes, 8 November 2014. Four females from La Posta ( 26°00′46.21′′N, 97°58′59.00′′W, ca. 22 m asl), 2.2 km N of Buenavista, Rio Bravo, Tamaulipas, Mexico. Lichen on mesquite tree ( Prosopis glandulosa ) coll. Antonio Moreno - Talamantes, 19 December 2016.
Species description. Body white or transparent with light yellow to brownish tones before fixation ( Fig. 1a View FIGURE 1 ). Eyes present in all living and mounted adults. Hatchling and juveniles without eyes. Adults cuticle sculptured with pseudopores (0.6 – 1.4 μm in diameter), sparsely distributed and not forming a reticular design. Hatchling and juve- niles with reticular design ( Figs 1b View FIGURE 1 and 2b View FIGURE 2 ). In adults, dorsal side with delineated geometric areas i.e. pseudoplates clearly visible only in FM ( Figs 3 – 4 View FIGURE 3 View FIGURE 4 ). The row I of pseudoplates situated anteriorly to legs I with four (two small and two large) pseudoplates. The row II situated in line with legs I with two triangular pseudoplates connected in the middle. The row III situated between legs I and II with four connected rectangular pseudoplates forming a large rectangle in the midline of the body. The row IV situated in the line of legs II with six rectangular pseudoplates forming a large rectangle in the midline of the body and two pairs of small oval dorsolateral pseudoplates. The row V situated between legs II and III with two rectangular pseudoplates connected in the middle, and two pairs of small oval dorsolateral pseudoplates. The row VI situated in line with legs III with six rectangular pseudoplates forming a large rectangle in the midline of the body and two pairs of small oval dorsolateral pseudoplates. The rows VII to IX situated between legs III and IV (row VII just behind legs III, row VIII in the middle between legs III and IV, row IX just before legs IV). The row VII with a double rectangular paired pseudoplate (the caudal pair poorly developed) and two pairs of small oval dorsolateral pseudoplates. The row VIII with a complex of ten pseudoplates, eight clustered on the dorsal side and two oval placed dorso - laterally. The row IX with a pair of pseudoplates connected in the middle. The pseudoplates arrangements see Remarks below. For pseudoplates in row I and II poorly visible under PCM, but clearly visible in FM. ( Fig. 4 View FIGURE 4 ). The pseudoplates in the hatchlings and juveniles poorly developed.
Buccal apparatus of the Milnesium type ( Figs 1c View FIGURE 1 and 2c View FIGURE 2 ). Buccal tube in adults wide and short (standard width on average 56% of its length) and funnel - shaped, wider anteriorly (posterior diameter on average 89% of the anterior diameter); buccal tube in hatchling and juveniles is more slender (standard width on average 35% of its length) and funnel - shaped, wider anteriorly (posterior diameter on average 80% of the anterior diameter). Six peribuccal papillae and six peribuccal lamellae (of unequal size, 4+2) around the mouth opening present. Two cephalic papillae positioned laterally. The peribuccal papillae longer than the cephalic one. Pharyngeal bulb elongated and pear - shaped, without placoids or septulum.
Claws of the Milnesiidae type ( Figs 1 View FIGURE 1 d–e and 2d–e). Primary branches on all claws with small accessory points. Secondary branches of all claws with rounded basal thickenings. Transverse cuticular bars present under claws I – III. In adult specimens, internal secondary branches on legs I–III with three points, external with two points. Secondary branches of anterior claw on legs IV with three points, posterior claws with two points. In hatchling and juvenile specimens all secondary branches on all legs with two points. Milnesium cassandrae sp. nov. exhibits an early positive internal CC (claw configuration [2 - 2/3] - [2/3 - 2]); in other words, changes in claw configuration ( CC) occur between the hatchling/juvenile stages [2 - 2] - [2 - 2], and adults [2 - 3] - [3 - 2].
Males: not observed.
Eggs: Oval, smooth and deposited in exuvium as in all other known Milnesium species.
Type depositories: The holotype (slide ELITE - 1/053) and 11 paratypes (slide: ELITE - 1/052, ELITE - 2/054 - 055 - 056 - 057, ELITE - 4/089, ELITE - 5/091 - 092 - 093, ELITE - 30/ 1068 - 1069), one exuviae with four eggs (slide ELITE - 30/1067) are deposited at Colección Carcinológica - FCB - UANL at Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, San Nicolás de los Garza, Nuevo León, Mexico, nine paratypes (slides ELITE - 12/878, ELITE - 14/881, ELITE - 43/ 1097 - 1098 - 1099, ELITE - 44/ 1100 - 1101 - 1102, ELITE - 52/1115), one exuviae and eight eggs ( ELITE - 52/1114) are preserved at the Department of Animal Taxonomy and Ecology, Adam Mickiewicz University, Poznań, Uniwersytetu Poznańskiego 6, Poznań, Poland and 27 paratypes (slides: ELITE - 3/058 - 060, ELITE - 13/879 - 880, ELITE - 28/1060, ELITE - 29/ 1065 - 1066, ELITE - 35/ 1084 - 1085, ELITE - 36/ 1086 - 1087, ELITE - 37/1089, ELITE - 40/1092, ELITE - 41/1094, ELITE - 42/1096. ELITE - 45/1103, ELITE - 47/1106, ELITE - 49/ 1108 - 1109, ELITE - 50/ 1110 - 1111, ELITE - 53/1116, ELITE - 54/ 1119 - 1120, ELITE - 58/1139, ELITE - 59/ 1140 - 1141), eight exuviae with 13 eggs (slides: ELITE - 27/987, ELITE - 29/1064, ELITE - 36/1088, ELITE - 38/1090, ELITE - 39/1091, ELITE - 53/1117, ELITE - 54/1118) are deposited in the collection of first author.
Etymology: The new species is dedicated to Cassandra Moreno Perales, the first author’s youngest daughter.
Remarks: To clarify the nomenclature of the pseudoplates arrangement we propose the system which is similar to that used for the numbering of gibbosities in Doryphoribius and Isohypsibius species by Michalczyk & Kaczmarek (2010). In this system all rows of pseudoplates are named and assigned to a specific place on the dorsal cuticle and the number of pseudoplates is counted:
row I of pseudoplates situated anteriorly to legs I,
row II of pseudoplates situated in line with legs I,
row III situated between legs I and II,
row IV situated in line with legs II,
row V situated between legs II and III,
row VI situated in line with legs III,
row VII situated between legs III and IV (just behind legs III),
row VIII situated between legs III and IV (in the middle between legs III and IV),
row IX situated between legs III and IV (just before legs IV).
The row numbers are always in Roman numerals and the number of pseudoplates is always listed in Arabic numerals. For example, in this system a configuration of pseudoplates (CP) in Mil. cassandrae sp. nov. will be: (CP: I:4; II:2; III:4; IV:10; V:6; VI:10; VII:8; VIII:10; IX:2, Figs 3 – 4 View FIGURE 3 View FIGURE 4 ).
Differential diagnosis. Based on having a sculptured dorsal cuticle, Mil. cassandrae sp. nov. belongs to the granulatum group ( Michalczyk et al. 2012a, b). The new species with claw configuration [2 - 3] - [3 - 2] in adults, is most similar to Mil. krzysztofi Kaczmarek & Michalczyk, 2007 and Mil. beasleyi Kaczmarek et al., 2012 , but it differs from:
1. Milnesium krzysztofi by: different dorsal sculpture (very dense pseudopores, but not forming a reticular design in adults of Mil. cassandrae sp. nov. vs. a fine reticular design in Mil. krzysztofi ), presence of eyes, a higher pt of the standard buccal tube width ([ 41.6–67.2] in Mil. cassandrae sp. nov. and [ 33.1–38.4] in Mil. krzysztofi ), and the presence of pseudoplates (but see Discussion, below).
2. Milnesium beasleyi by: presence of eyes, having higher pt buccal tube width ([ 47.4–66.2], [ 41.6–67.2], [ 39.9– 61.1] anterior, standard and posterior, respectively in Mil. cassandrae sp. nov. vs. [ 35.3–41.8], [ 31.2–39.8], [ 33.2–39.6] anterior, standard and posterior, respectively in Mil. beasleyi ), higher standard width/ length ratio of buccal tube (42% – 67% in Mil. cassandrae sp. nov. vs. 31% – 40% in Mil. beasleyi ), smaller lateral papillae (4.0 – 6.5 in Mil. cassandrae sp. nov. vs. 7.5 – 10.3 in Mil. beasleyi ), and presence of pseudoplates (but see Discussion, below).
In addition, Mil. cassandrae sp. nov. is similar to other species with the claw configuration [2 - 3] - [3 - 2] i.e. Mil. lagniappe Meyer et al., 2013 , Mil. reductum Tumanov, 2006 , Mil. reticulatum Pilato et al., 2002 , Mil. tardigradum tardigradum Doyère, 1840 , Mil. tetralamellatum Pilato & Binda, 1991 and Mil. vorax Pilato et al., 2016 , but it differs from:
3. Milnesium lagniappe by: larger number of peribuccal lamellae (six in Mil. cassandrae sp. nov. vs. four in Mil. lagniappe ), different dorsal sculpture (very dense pseudopores, but not forming a reticular design adults of Mil. cassandrae sp. nov. vs. reticulated pattern of irregular polygons in Mil. lagniappe ), presence of eyes, stylet supports inserted in less caudal position ([ 58.7–67.6] in Mil. cassandrae sp. nov. vs. [ 69.7–73.4] in Mil. lagniappe ), and lower pt of buccal tube width ([ 47.4–66.2], [ 39.9–61.1] anterior and posterior, respectively, in Mil. cassandrae sp. nov. vs. [ 68.0–77.5], [ 61.8–70.8] anterior and posterior, respectively, in Mil. lagniappe ).
4. Milnesium reductum by: sculptured dorsal cuticle, presence of accessory points on primary branches of claws, stylet supports inserted in more anterior position (16.5 – 26.0 in Mil. cassandrae sp. nov. vs. 29.2 – 28.1 in Mil. reductum ), smaller claws IV (see Table 2 View TABLE 2 in this work and Table 5 in Tumanov (2006) for the exact differences in dimensions of claws), and presence of pseudoplates (but see Discussion, below).
5. Milnesium reticulatum by: larger number of peribuccal lamellae (six in Mil. cassandrae sp. nov. vs. four in Mil. reticulatum ), absence of cuticular gibbosities, different dorsal sculpture (very dense pseudopores, but not forming a reticular design in adults of Mil. cassandrae sp. nov. vs. pseudopores arranged in nine sculptured bands, forming a reticular design in Mil. reticulatum ), higher pt of the standard buccal tube width ([ 45.6–67.2] in Mil. cassandrae sp. nov. vs. [ 30.4–37.4] in Mil. reticulatum ), and higher pt of claws II, III and IV (see Table 2 View TABLE 2 in this work and Table 2 View TABLE 2 in Pilato et al. (2002) for the exact differences in dimensions of claws).
6. Milnesium tardigradum tardigradum by: having sculptured dorsal cuticle and presence of pseudoplates (but see Discussion, below).
7. Milnesium tetralamellatum by: larger number of peribuccal lamellae (six in Mil. cassandrae sp. nov. vs. four in Mil. tetralamellatum ), sculptured dorsal cuticle, and presence of pseudoplates (but see Discussion, below).
8. Milnesium vorax by: sculptured dorsal cuticle, longer % external base + secondary branch/external primary branch I to III in Mil. cassandrae sp. nov. (see Table 2 View TABLE 2 in this work and Table 1 View TABLE 1 in Pilato et al. (2016) for the exact differences), and presence of pseudoplates (but see Discussion, below).
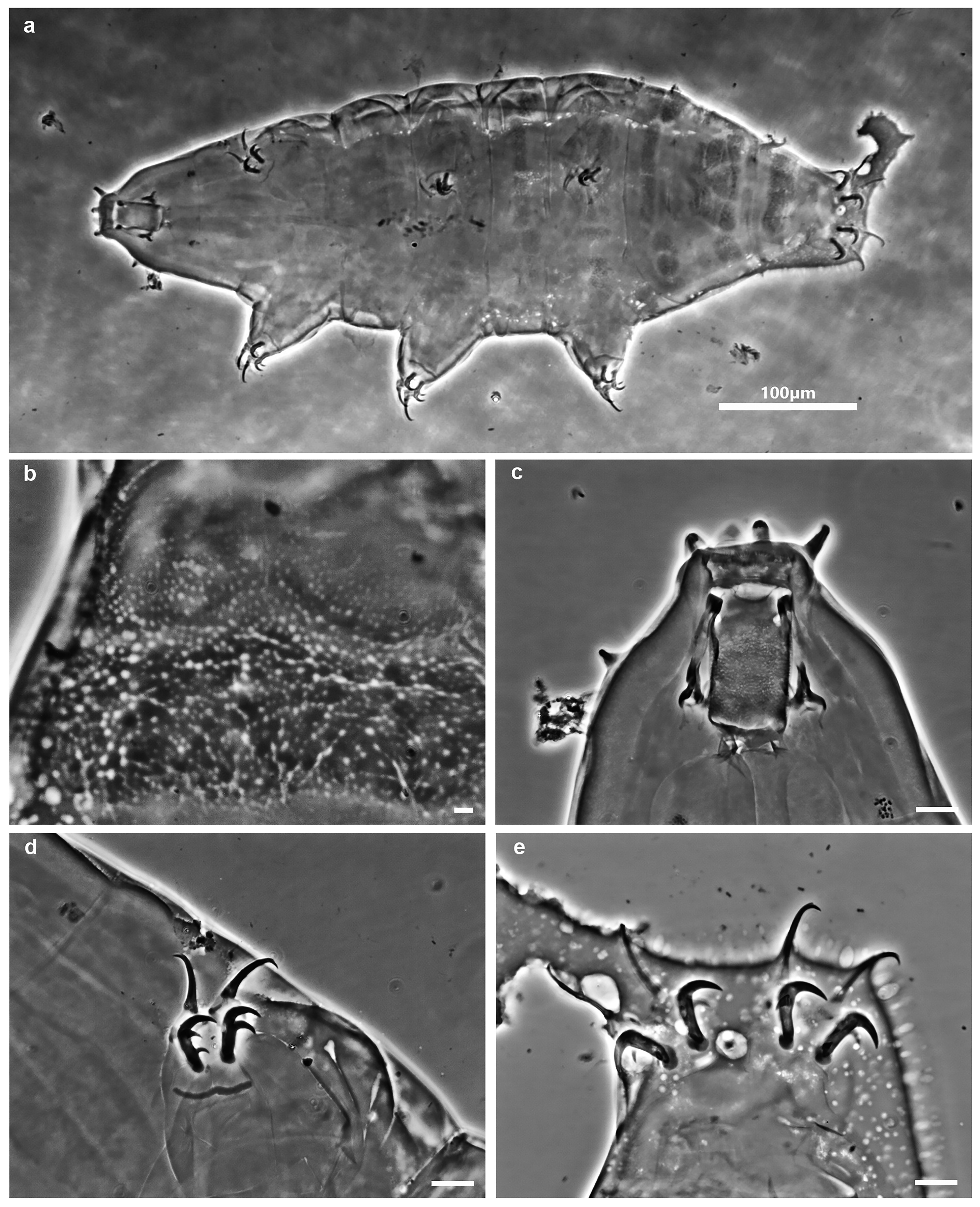
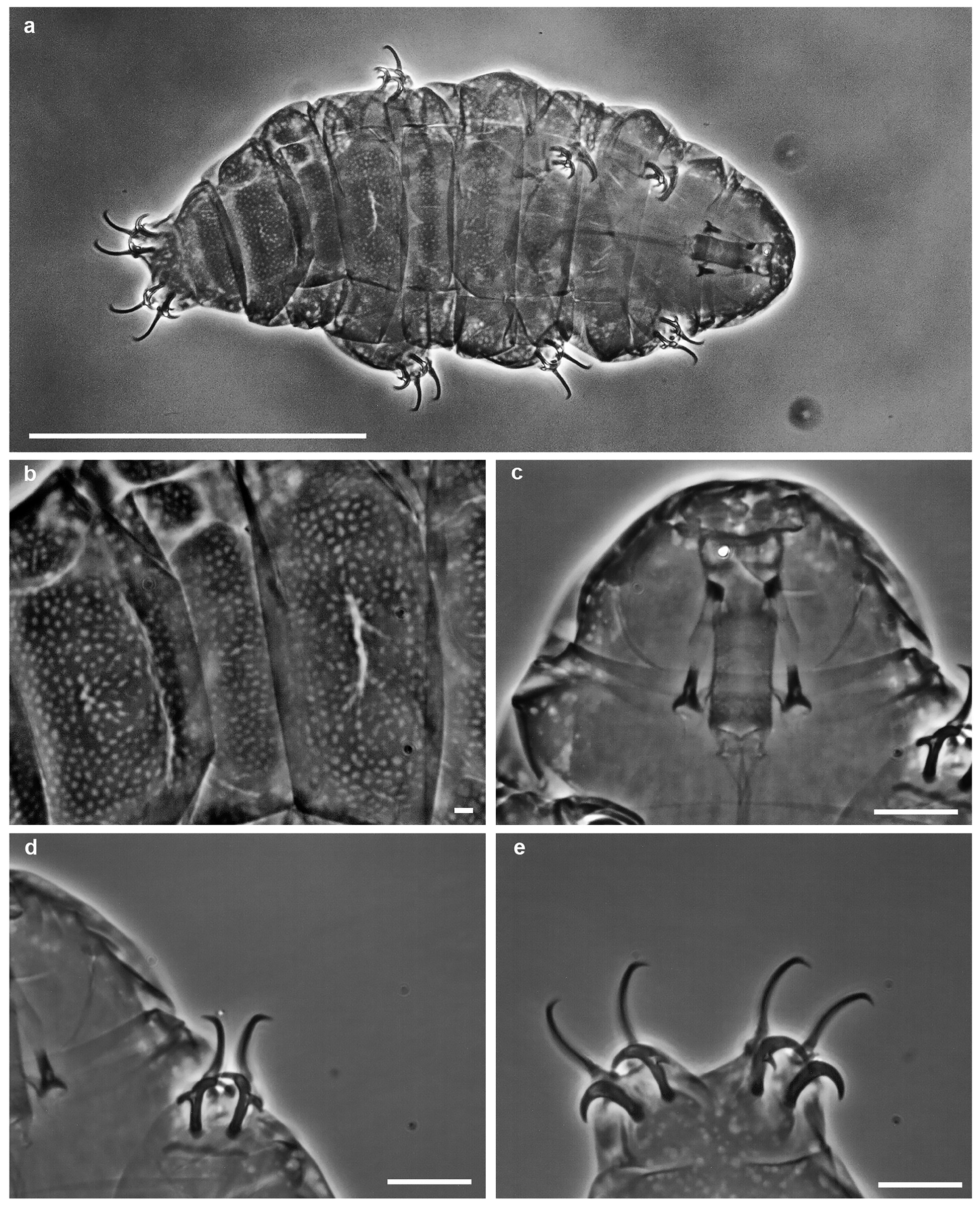

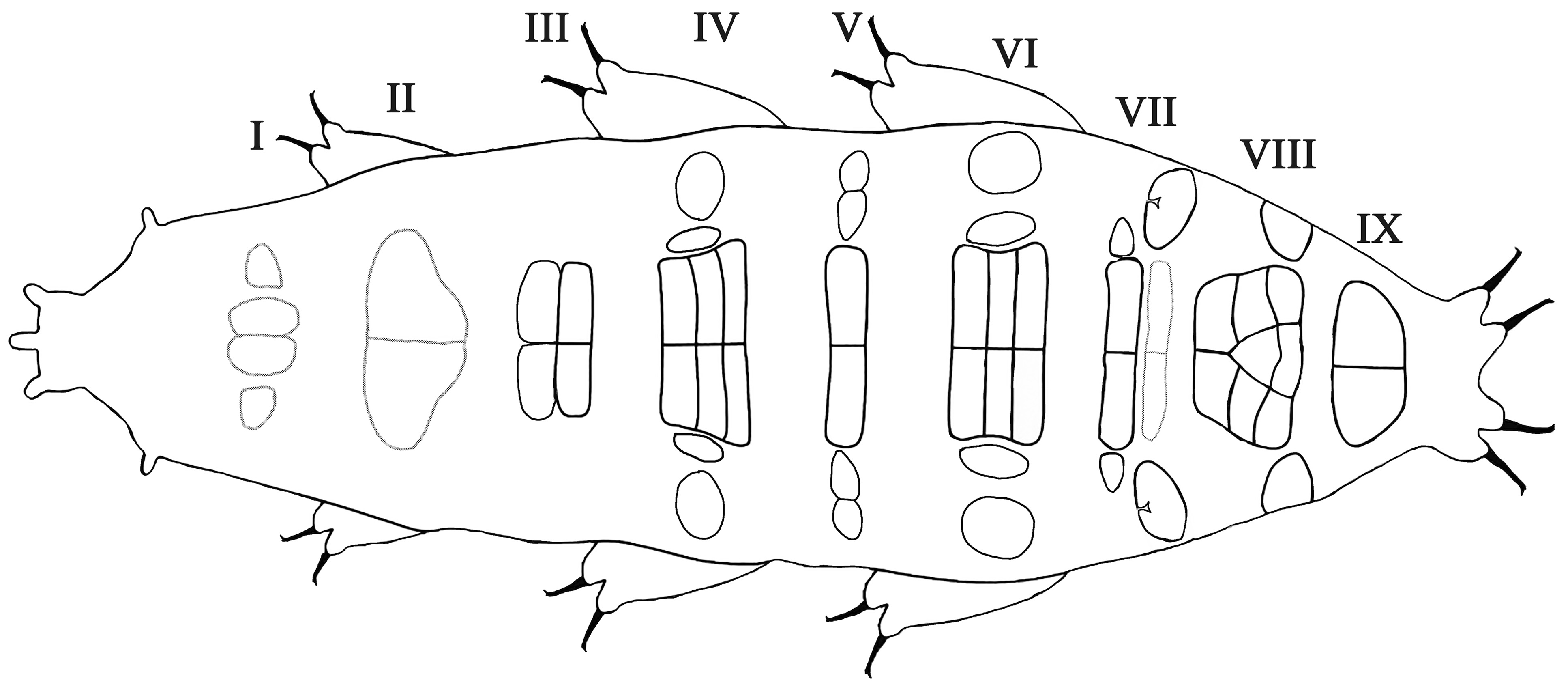
TABLE 3. Measurements and pt values of selected morphological structures of 13 hatchling and juvenile specimens (instar I and II) of Milnesium cassandrae sp. nov. mounted in PVA medium. Range refers to the smallest and the largest structure among all measured specimens. The pt values are provided in italics; N, number of specimens or structures measured; SD, standard deviation.
| Character | N | Range µm | pt | Mean µm | pt | SD µm | pt | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Body length | 13 | 131 | – | 202 | – | 179 | 20 | ||||
| Peribuccal papillae length | 3 | 3.3 | – | 4.2 | 16.2 | – | 19.7 | 3.7 | 17.9 | 0.4 | 1.7 |
| Lateral papillae length | 5 | 2.3 | – | 3.4 | 11.3 | – | 16.5 | 3.0 | 14.6 | 0.4 | 2.1 |
| Buccal tube | |||||||||||
| Length | 13 | 18.2 | – | 22.6 | – | 20.7 | – | 1.2 | – | ||
| Stylet support insertion point | 13 | 10.3 | – | 14.8 | 56.5 | – | 70.5 | 13.8 | 66.3 | 1.1 | 3.4 |
| Anterior width | 13 | 7.2 | – | 9.6 | 38.1 | – | 47.1 | 8.8 | 42.3 | 0.7 | 3.2 |
| Standard width | 13 | 6.1 | – | 7.8 | 28.2 | – | 39.4 | 7.2 | 34.6 | 0.5 | 3.0 |
| Posterior width | 13 | 6.3 | – | 7.3 | 29.6 | – | 36.1 | 7.0 | 33.6 | 0.3 | 2.0 |
| Standard width/length ratio | 13 | 28% | – | 39% | – | 35% | – | 3% | – | ||
| Posterior/anterior width ratio | 13 | 74% | – | 88% | – | 80% | – | 4% | – | ||
| Claw 1 lengths | |||||||||||
| External primary branch | 13 | 7.4 | – | 10.4 | 35.1 | – | 52.8 | 8.9 | 42.9 | 0.9 | 5.7 |
| External base + secondary branch | 12 | 6.5 | – | 7.9 | 30.1 | – | 39.7 | 7.3 | 35.1 | 0.4 | 2.6 |
| External branches length ratio | 12 | 72% | – | 96% | – | 84% | – | 10% | – | ||
| Internal primary branch | 13 | 7.6 | – | 9.8 | 35.8 | – | 48.4 | 8.7 | 42.1 | 0.7 | 4.1 |
| Internal base + secondary branch | 13 | 6.2 | – | 8.2 | 30.2 | – | 39.6 | 7.1 | 34.3 | 0.5 | 2.9 |
| Internal branches length ratio | 13 | 74% | – | 94% | – | 82% | – | 6% | – | ||
| Claw 2 lengths | |||||||||||
| External primary branch | 13 | 7.6 | – | 11.0 | 35.4 | – | 60.5 | 9.3 | 45.2 | 0.9 | 6.5 |
| External base + secondary branch | 13 | 6.7 | – | 8.1 | 30.9 | – | 40.9 | 7.5 | 36.3 | 0.5 | 3.5 |
| External branches length ratio | 13 | 65% | – | 94% | – | 81% | – | 7% | – | ||
| Internal primary branch | 12 | 7.6 | – | 9.7 | 37.4 | – | 53.5 | 9.0 | 43.3 | 0.6 | 4.3 |
| Internal base + secondary branch | 13 | 6.4 | – | 8.9 | 29.9 | – | 40.9 | 7.2 | 34.7 | 0.7 | 3.4 |
| Internal spur | 1 | 1.8 | – | 1.8 | 10.1 | – | 10.1 | 1.8 | 10.1 | ? | ? |
| Internal branches length ratio | 12 | 70% | – | 96% | – | 80% | – | 9% | – | ||
| Claw 3 lengths | |||||||||||
| External primary branch | 12 | 8.5 | – | 10.3 | 39.4 | – | 48.9 | 9.4 | 44.7 | 0.5 | 3.2 |
| External base + secondary branch | 13 | 6.0 | – | 8.3 | 31.3 | – | 38.5 | 7.3 | 35.1 | 0.6 | 2.3 |
| External branches length ratio | 12 | 71% | – | 90% | – | 79% | – | 6% | – | ||
| Internal primary branch | 12 | 8.6 | – | 9.7 | 39.0 | – | 47.2 | 9.3 | 44.4 | 0.4 | 2.8 |
| Internal base + secondary branch | 13 | 6.7 | – | 7.6 | 30.4 | – | 40.0 | 7.1 | 34.2 | 0.3 | 2.7 |
| Internal branches length ratio | 12 | 70% | – | 83% | – | 76% | – | 4% | – | ||
| Claw 4 lengths | |||||||||||
| Anterior primary branch | 11 | 8.4 | – | 11.2 | 41.5 | – | 54.9 | 10.2 | 49.7 | 0.8 | 3.6 |
| Anterior base + secondary branch | 13 | 5.9 | – | 8.1 | 26.1 | – | 40.7 | 7.1 | 34.4 | 0.5 | 3.7 |
| Anterior branches length ratio | 11 | 61% | – | 80% | – | 71% | – | 6% | – | ||
| Posterior primary branch | 11 | 9.5 | – | 11.8 | 45.8 | – | 56.7 | 10.7 | 52.3 | 0.7 | 3.3 |
| Posterior base + secondary branch | 13 | 6.6 | – | 8.2 | 30.6 | – | 40.9 | 7.5 | 36.1 | 0.5 | 3.1 |
| Posterior branches length ratio | 11 | 65% | – | 77% | – | 70% | – | 4% | – | ||
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |