
Malo maxima, Gershwin, Lisa-Ann, 2005
|
publication ID |
https://doi.org/ 10.5281/zenodo.170452 |
|
DOI |
https://doi.org/10.5281/zenodo.6269009 |
|
persistent identifier |
https://treatment.plazi.org/id/03E9878B-AC58-3462-FE95-CA54D791F9B8 |
|
treatment provided by |
Plazi |
|
scientific name |
Malo maxima |
| status |
sp. nov. |
Malo maxima sp. nov.
Figures 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 , Figure 6 View FIGURE 6 D–F
Material examined
Holotype: WAM Z29940 (=B54), offshore from 80 Mile Beach, in the pearling grounds South of Broome, Kimberley coast, Western Australia [approx. 19° 13.529S 121° 15.856E], 15 May 2004, caught by nightlight aboard the Paspaley Clare II; live measurements: 48mm BH, 40.5mm DBW, 20.5mm IRW; preserved measurements: 43.20mm BH, 36.66mm DBW, 17.97mm IRW, 1.46mm TBW, 2.97mm VW, 19.69mm PL, 8.38mm PW, 1.47mm PCW, 2.54mm OKW, 4.62mm IKW.
Paratypes: WAM Z29941, same locality as holotype, 16 April 2004, caught aboard the Paspaley Marilynne; gravid female, 27.50mm BH, 28.18mm DBW, 13.09mm IRW, 1.81mm TBW. WAM Z29946, 10km off Gantheaume Point, Broome, 13 July 2004, caught by nightlight by the crew of the Paspaley Marilynne; lot of 14, 36.62mm BH to 23.16mm BH.
Paratypes (all paratypes in this paragraph from same general locality as holotype, caught by nightlight aboard the Paspaley Clare II): WAM Z29942 (=B26), male, 22.72mm BH, 20.45mm DBW, 10.05mm IRW, 1.37mm TBW. WAM Z29943 (=B3), male, 28.37mm BH, 27.43mm DBW, 12.80mm IRW, 1.56mm TBW. WAM Z29944 (=B4), male, 25.73mm BH, 25.86mm DBW, 12.64mm IRW, 1.37mm TBW. WAM Z29945 (=B8), male, 26.59mm BH, 25.18mm DBW, 12.58mm IRW, 1.67mm TBW. WAM Z29947 (=B7), immature male, 25.68mm BH, 23.41mm DBW, 11.21mm IRW, 1.70mm TBW. WAM Z29948 (=B42), immature male, 22.93mm BH, 24.66mm DBW, 12.09mm IRW, 1.43mm TBW. WAM Z29949 (=B44), immature female, 22.00mm BH, 21.71mm DBW, 10.65mm IRW, 1.68mm TBW. WAM Z29942 (=B55), 17mm BH in life. QM G324167 (=B5), male, 35.00mm BH, 32.20mm DBW, 15.68mm IRW, 1.20mm TBW. AM G17010 (=B6), male, 31.47mm BH, 29.89mm DBW, 15.18mm IRW, 1.30mm TBW. SAM H1409 (=B2), male, 34.29mm BH, 31.00mm DBW, 15.35mm IRW, 1.23mm TBW. NTM C15023 (=B31), gravid female, 32.80mm BH, 29.01mm DBW, 12.97mm IRW, 1.37mm TBW. MTQ G55400 (=B32), male, 32.18mm BH, 32.08mm DBW, 16.35mm IRW, 1.68mm TBW.
Other material: Unregistered specimen in the collection of Menzies School of Health Research, Royal Darwin Hospital, Darwin, N.T., coll. 19 May 2002 by the Paspaley Clare II crew; male, 32.79mm BH, 30.25mm DBW, 15.35mm IRW, 1.33mm TBW, 17.56mm PL, 5.81mm PW, 1.33 PCW, 2.45mm IKW, 1.96mm OKW. Approximately 100 specimens, same collection data as holotype, examined casually, photographed, and measured while alive, then sacrificed for venom research.
Type locality
Off 80 Mile Beach, south of Broome, Kimberley coast, Western Australia.
Etymology
The species name is a noun in apposition, derived from the name of the pearl oyster, Pinctada maxima , and was chosen by the Pearl Producers Association to honour the pearl divers of Western Australia, who are the most commonly and severely affected by the presumed sting of this species.
Diagnosis
Malo with tall, narrow, robust body, to about 5cm BH; with flattened apex; with small, narrow pedalia, each with a single row of nematocyst patches on outer keel; rhopalial horns short, broad; 45 velarial canals per octant, simple to bifurcated, arising from single wide base at velarial turnover; with flaplike perradial mesenteries to about 1/3 BH toward rhopalia.
Description of holotype
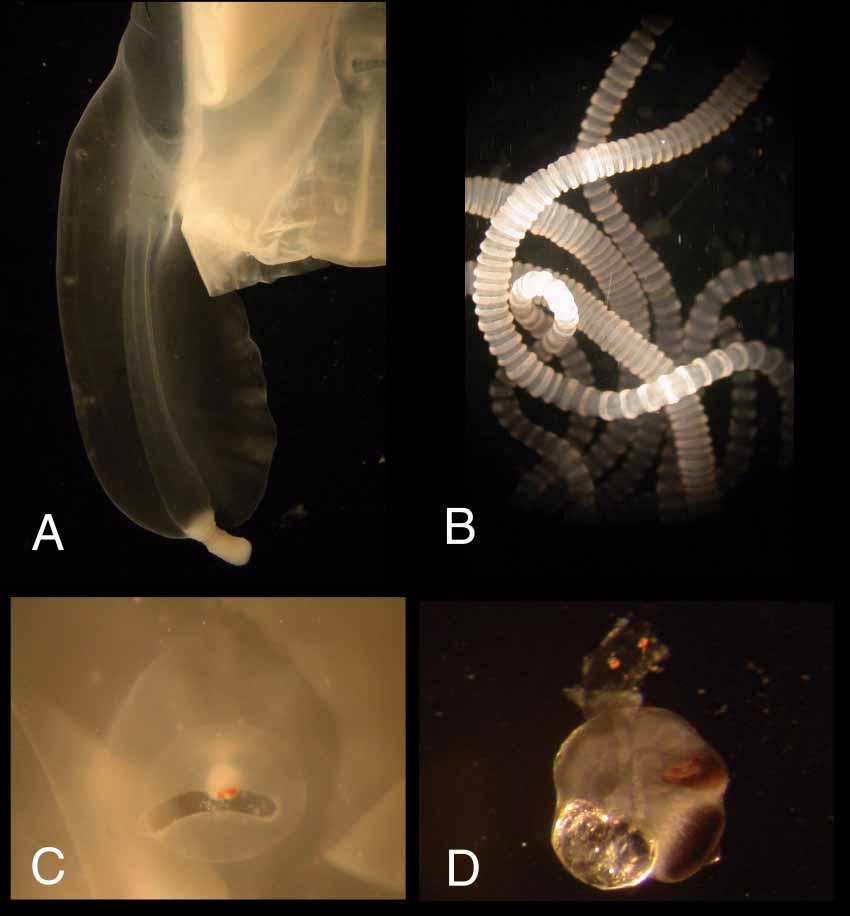
Body ( Figure 2 View FIGURE 2 ) of medium size, about 5 cm tall in life, more than twice as tall as wide; robust, evenly thick, keeping its shape in water. Apex conspicuously flattened, fairly densely covered with gelatinous nematocyst warts; lacking reticulated pattern or coronal furrow. Conspicuous gelatinous nematocyst warts additionally scattered over entire exumbrella. Interradial furrows deep, from level with stomach down to top of pedalia. Adradial furrows well pronounced in upper half of body, defining interradial pillars; well demarcating perradial rhopaliar region from pillars in lower half of body.
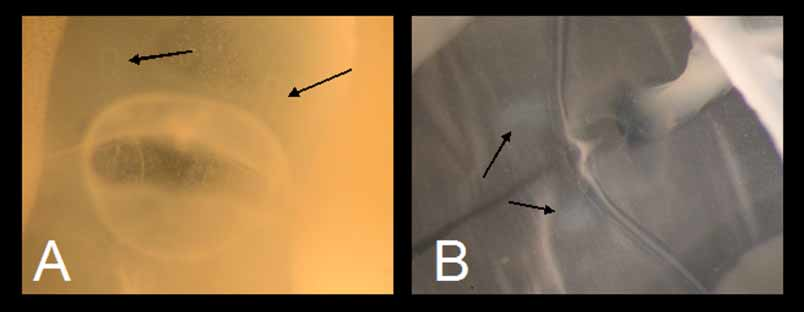
Pedalia 4, interradial, one per corner, simple, unbranched, approximately 1/2 BH measured from subumbrellar lamella, but only hanging down past velarium about 1/4 as long as bell is tall; with three small unraised nematocyst patches (“freckles” rather than “warts”) evenly spaced in upper half of outer wing ( Figure 3 View FIGURE 3 A). Inner keel overhangs at point of tentacle insertion. Pedalium narrow scalpelshaped, with inner keel about 2 1/ 2x width of canal, outer keel about 1 1/ 2x as wide as canal ( Figure 3 View FIGURE 3 A). Pedalial canals substantially quadrate in cross section throughout, with a short, blunt, obliquelypointing projection at bend near base; straight to slightly flared at point of tentacle insertion ( Figure 3 View FIGURE 3 A). Tentacles 4, 1 per pedalium, unbranched; fine, round in cross section ( Figure 3 View FIGURE 3 B); base straightsided, about 1.46mm diameter; banding of contracted tentacles alternating smaller with larger (i.e., 121212), in a few places appearing evenly banded (i.e., 111 111); with a macrosegmented appearance of repeating hourglass pattern when contracted.
Rhopalial niche prominently raised from bell wall, plus entire rhopalial area raised up from body wall by oblique portions of deeplycut adradial furrows. Rhopalial niche ostium frownshaped, with a single, smoothedged covering scale each above and below; upper scale shallowly concave, lower scale shallowly convex ( Figure 3 View FIGURE 3 C). Rhopalial horns short, thick, straight to somewhat undulating ( Figure 4 View FIGURE 4 A, B, arrows). Rhopalial warts not observed. Eyes 2 per rhopalium, median with lenses; lateral eye spots lacking ( Figure 3 View FIGURE 3 D).
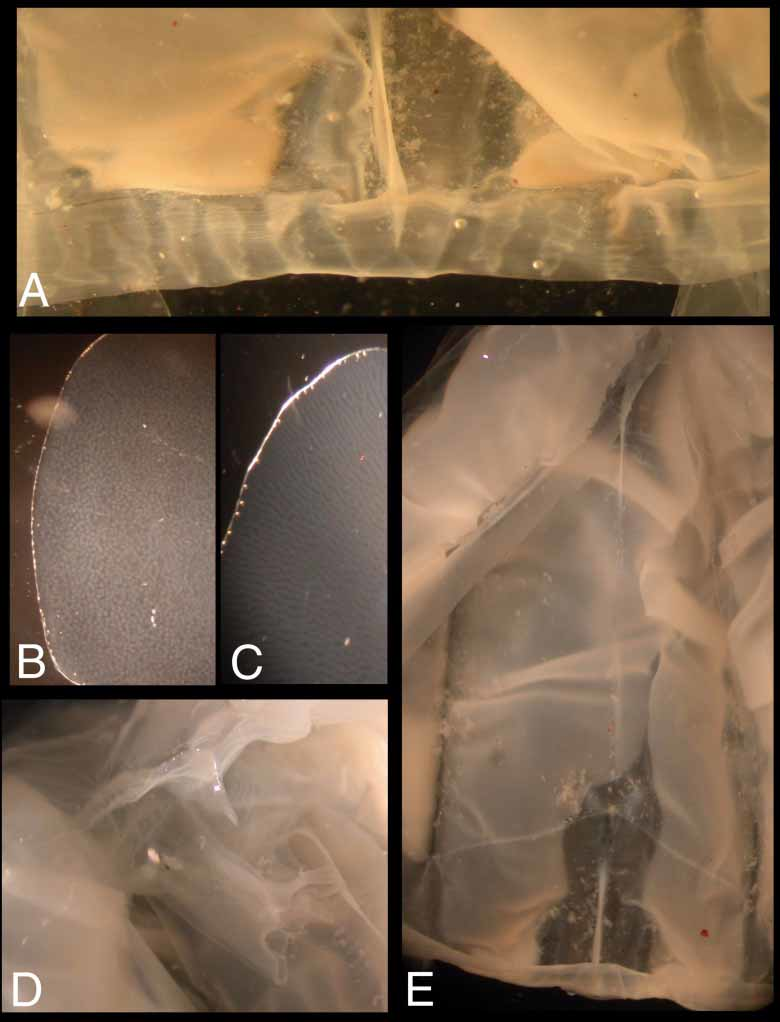
Velarium ( Figure 5 View FIGURE 5 A) quite narrow, with a single wide canal coming from stomach pouch in each octant, branching at velarial turnover into approximately 4 simple to bifurcated canals with undulating edges; 46 velarial canal tips reaching margin or nearly so. Perradial lappets 4, narrow, stopping short of velarial margin; each side with a conspicuous, large, gelatinous, raised nematocyst wart, not corresponding in position; extended distally by nipplelike canals; lacking lateral canals ( Figure 5 View FIGURE 5 A). Frenulum a single gelatinous sheet, extending from lower border of rhopaliar niche to reaching about threequarter distance toward velarial margin ( Figure 5 View FIGURE 5 A, E).
Phacellae absent. Lateral gonads leaflike, attached along entire height of interradial septa; overlapping perradially, pleated, narrowed in lower 1/3 of bell around rhopaliar niche; extending down through radial pouches into bases of velarial canals. Gonads of holotype with “fingerprint” pattern characteristic of male cubozoan gonads ( Figure 5 View FIGURE 5 B), lacking eggs characteristic of female gonads ( Figure 5 View FIGURE 5 C). Interradial septum perforated.
Stomach quadrate in cross section; short and flat in preserved specimen, large and baglike in life. Proboscis very small and very short ( Figure 5 View FIGURE 5 D), 7.11mm long to tips of short, somewhat crenulated lips (preserved). Perradial mesenteries moderately well developed, flaplike in upper 1/3 bell height, with sessile band of thickened tissue extending to rhopalial stem ( Figure 5 View FIGURE 5 E). Subumbrellar rhopalial windows smooth and flat, with a small concavity at top where rhopalium joins nerve cord ( Figure 5 View FIGURE 5 E).
Colour: in life, body and gonads transparent and colourless, with a hint of cloudiness to mesoglea; tentacles pale pink and about 5– 8 x BH in normal swimming posture, exumbrellar rhopaliar warts pale purple, rhopalia appeared as small black spots; after two months preserved in dilute formalin (equal to approx. 2% formaldehyde), gonads and tentacles opaque whitish, mesoglea hyaline, nematocyst warts faded to nearly invisible, and eye spots faded brown.
Variation
Most of the paratypes have the following characters which are different to the holotype, presumed to be related to growth: the rhopalial horns are curved inward slightly, resembling devil horns or Viking horns, but in all cases they are still short and thick; the perradial lappets typically have a row of 1–4 nematocyst warts on each side (typically 2), almost always mirrored between the opposing lappets in each pair; the velarial canals are typically branched into 4 or 5 fingers, which may or may not have side branches; the perradial lappets rarely have side branches, but almost always have terminal nipplelike extensions; the pedalia typically have between 1–10 nematocyst warts in a row on the outer keel, most often about 3–4; the pedalial canal bend is highly variable from kneelike, to 90°cornered, to having a very low upwardpointing thorn where the canal breaks away from the lamella, to resembling that described for the holotype, i.e., with a blunt obliqueedged extension. All other characters were consistent with those described for the holotype.
Specimen number WAM Z29948 has two halflength pedalia on one of the corners, for a total of 5 pedalia; the remaining three pedalia are of the normal length. Specimen number WAM Z29945 has a crenulated margin of the inner pedalial keels, and in specimen number WAM Z29948, the margin is instead notched and lobed. All of these are believed to be mere aberrations.
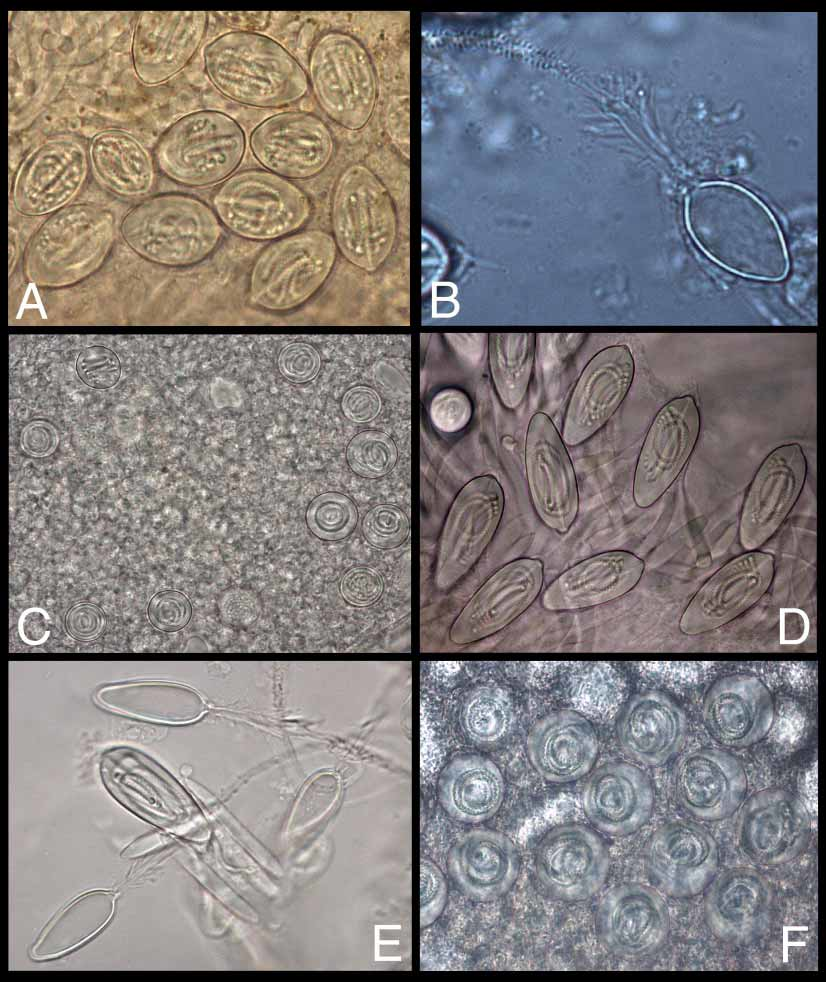
Nematocysts ( Figure 6 View FIGURE 6 D–F)
The tentacular nematocysts of M. maxima are of one type only, namely penetrantform microbasic pmastigophores with all spines orientated away from the capsule ( Figure 6 View FIGURE 6 D, E). The nematocysts have an arithmetic mean of 43.18µm long by 16.87µm at the widest point (range 34.55–49.32µm long by 14.59–19.65µm wide, n=58). The capsule is exaggeratedly elongate, resembling a cross between a cigar and an egg, wider toward the distal end and with a distinct nipple at the distal apex; the proximal end narrows to a smoothly rounded point. Inside the capsule, a baseballbatshaped shaft extends about 2/3 the length, wider at its distal end (= the proximal end of the capsule), with the spined tubule coiled loosely five times in the longitudinal axis. The discharged basis is evenly cylindrical, with distallypointing spines along the entire length; the basis is about half again as wide as the tubule, with a tapered joint. The tubule is of an even diameter and evenly armed with several spiraling rows of small spines throughout. Many of the observed capsules that were no longer embedded in the tentacle lacked spines, suggesting that the spines may fall off easily, especially when they have discharged into an object (in this case, probably another portion of the tentacle, or maybe the collecting container). The lack of spines could prove confusing in nematocyst identification, particularly in medical diagnosis of Irukandji patients; however, the capsule shape and size, as well as the shaft form are still diagnostic.
Three types of microbasic pmastigophores are currently differentiated based on direction of spination: Type 1, with all spines orientated at right angles to the capsule; Type 2, with all spines orientated toward the capsule; and Type 3, with most spines short and pointing toward the capsule, plus some longer spines pointing away ( Williamson et al. 1996). The nematocysts of Malo maxima have only large spines, which all point away from the capsule ( Figure 6 View FIGURE 6 E), herein designated as Type 4; this same arrangement is also found in at least five other closelyrelated undescribed species, namely “Morbakka,” the “Darwin carybdeid,” “PseudoIrukandji,” “HaloIrukandji,” and the “Dampier Irukandji.” This type of nematocyst is similar to the form erroneously attributed by Wiltshire et al. (2000) to Carukia barnesi ; neither the specimen (fig. 2) nor the nematocysts (fig. 3) match those of C. barnesi ( Southcott 1967) .
The capsule shape of the mastigophores of Malo maxima is reminiscent of that of Chironex fleckeri and many of the chirodropids in that both forms are elongate, but these are considerably more eggshaped and relatively wider than those of the chirodropids. In comparison with the discharged basis and tubule of the common carybdeid microbasic euryteles (= basis dilated distally, with numerous rows of relatively small spines) and tumiteles (= basis dilated midway, with numerous large spines), these bases are longer and cylindrical and the spines are prominent throughout the length. One would not easily confuse these nematocysts with the typical carybdeid nematocysts, or with those of the chirodropids.
This brings up an issue that has plagued recent cubozoologists ( Kingston and Southcott 1960; Southcott 1967; Calder and Peters 1975; Williamson et al. 1996; Yanagihara et al. 2002): most cubozoan nematocysts can be classified into one of the existing shaftbased groupings, but do not look like those of other cnidarian classes. For example, the principal tentacular nematocysts of many cubozoans are microbasic mastigophores (= cylindrical basis) by conventional interpretation, but the capsule shape, size of the spines, and pattern of spination are wholly different from those belonging to the Hydrozoa and Anthozoa. Thus, comparison of mastigophores between the Cubozoa and other classes ( Marques and Collins 2004) is potentially misleading, because the classification groups highly heterogeneous forms and may therefore be artificial. Similarly, the principal tentacular nematocysts of Carybdea and Carukia are microbasic euryteles (or tumiteles, used by some workers) based on the distal (or medial) swelling of the shaft, but again, the spines are completely different from those of the other classes. A revision of nematocyst taxonomy is badly needed, incorporating not only the basis and tubule morphology, but the forms of the capsules and spines as well.
The exumbrellar nematocysts of Malo maxima are large spherical isorhizas with a short, loosely coiled tubule, with an arithmetic mean of 28.32µm (range 23.59–29.82µm, n=49) ( Figure 6 View FIGURE 6 F). About 55–70 nematocysts were observed on each wart. The nematocysts of the pedalia, velarium, and lips were not examined.
Stinging ability
Malo maxima is apparently an extremely dangerous animal. I hypothesize that this is the species that left a fit, healthy male, 41 years old, in critical condition on life support for three days in February 2004, stung while surfing after a cyclone (which seems intriguing, because the previous life threatening sting, to a female in 2000, was also following a cyclone). This sting event progressed as follows (M. Longhurst, pers. comm., April 2004; his own words have been used as much as possible): The initial sting was felt as an immense sharp pain to the top of the back of the right thigh, under “board shorts” (i.e., surfer’s pants), like red hot pokers going into the leg. The initial pain lasted about ten minutes, with no welts or redness; it was not felt that repeated stinging took place, only a high level of pain that did not subside. The patient was aware of Irukandjis, but believed this was not to blame due to the severity of the pain of the sting. At about five minutes post sting, the patient felt an exertiontype pain in the region of the sternum. At about 10–15 minutes post sting, it became painful to breathe; pain was still localized to the sternum area. With breathing getting harder and shorter, the patient paddled back to the beach (about 15 minutes poststing). As soon as he stood up, he felt soreness in his back, which he attributed at the time to fatigue from not surfing for awhile. While walking the 300m across the beach, breathing was getting very difficult, the back was sore, and he noticed his legs getting sore, an achy feeling around the knees; he had to steady himself on the railing at the base of the stairs, feeling like he was losing control of his body and legs. Walking up the stairs, he was uncontrollably groaning and gasping quite loudly, such that people 100m away turned to look. The walk from the stairs to the car was an agonizing effort to force himself to walk, bent over, clutching his chest and surfboard. While trying to get into the car, he was unable to move his legs or use his arms to pull his legs in. He tried to phone for help but was unable to coordinate his movements enough to dial or hold the phone steady. At that point, the pain was not too bad, but the groaning was involuntary and the difficulty breathing was “freaking him out” (i.e., making him psychologically distressed). Some young men nearby came over to offer assistance, calling the hospital and transporting him in his car. During the tenminute drive to the hospital, he thought he would suffocate. At the hospital, he tried to get out of the car and fell in a heap on the ground, unable to move his legs or arms, laying there groaning and suffocating. Inside the hospital, his blood pressure was 230/110, and a total of 300mg of morphine still did not control the pain. He was intubated approximately five hours post sting, and transported by air ambulance to a hospital in Perth, where he remained unconscious for 2 1/2 days. In the following week, he experienced intense aching in calf muscles and the knees, with the knees feeling like they wanted to explode. Up to two weeks poststing, he felt sharp prickly sensations anywhere on the body, similar to the initial sting, as well as tightness in the chest and general fatigue. Some five months after the sting, he described a bout with the flu as “the worst flu ever,” and, while being laid out for three weeks, wondered if he had lost some tolerance from the syndrome.
Empirical evidence is lacking to support the hypothesis that Malo maxima was responsible for the sting of Mr. Longhurst. Skin scrapings of his sting were taken at the hospital in Broome, but I still await reply to my repeated requests for access to them. My hypothesis is not completely speculative, however, because a fatal sting in QLD with a similar severe initial sting, severe hypertension, and lack of nausea, has been attributed to an unknown species with very similar nematocysts ( Huynh et al. 2003). It is likely that the QLD fatality can be attributed to the undescribed “HaloIrukandji” or “PseudoIrukandji,” based on nematocysts recovered from the victim; this group was closely linked with M. maxima in a recent genetic study (Gershwin unpublished data).
Some 1012 severe syndromes occur per year that are treated on the pearling vessels (P. Cookson, Pearl Producers Association, pers. comm. 2004); some of these stings are apparently referable to M. maxima , as divers sometimes see themselves get stung by what they believe to be this species, subsequently developing the syndrome (various pearl divers and skippers, pers. comm. 2004). It is possible that the pearl divers have not had life threatening complications from severe envenomations because of their extreme level of athletic fitness. Countless other milder cases of Irukandji syndrome are experienced by the pearl divers, without narcotics being required. An additional 40 or so hospitalizations per year occur in Broome due to Irukandji syndrome (Kimberley Public Health Unit 2003; Macrokanis et al. 2004), though it is unclear for what percentage this species is responsible. This species has not yet proven to be lethal to humans, but all possible safety precautions should be used when handling or encountering this animal.
Distribution
So far this species is known only from off 80 Mile Beach and outside Roebuck Bay, Broome. It seems to be more common offshore; specimens have been caught nearshore during onshore winds, and several severe stings presumed to be from this species have occurred onshore following cyclones.
Ecology and natural history
The species is found in abundance offshore from the 80 Mile Beach region of North Western Australia, especially common at night when attracted by artificial lights. They are also said to often occur in a band about 9m deep, where they are commonly observed by the pearl divers during their decompression stops. They are only occasionally observed near the bottom, but this could be due to the divers’ diverted attention rather than due to species depth preferences.
For reasons that are not yet clear, the species was most often caught during the penultimate hour before slack tide, with greater abundance on a falling tide than on a rising tide. Approximately 1 1/2 to 2 hours before slack tide, medusae would begin coming to the stern of the boat, presumably drawn by powerful flood lights. They would increase in numbers, with often more than 10 visible at a time; they seemed undeterred in their attraction to the light by fishes and squids that were also swarming under the light. Abruptly about one hour to 1/2 hour before slack tide, the medusae would stop coming. Some nights they came for both slack tides but were more abundant on the earlier slack, while other nights they only came for the first slack tide. These results may have been biased by the fact that the collecting trip was limited to the neap period, during which the earlier evening tide was predominantly the low tide; it would be interesting to see what occurrence pattern the medusae exhibit during other periods of the tidal cycle. In contrast to the large number of medusae observed at night, approximately five specimens were caught and some several dozen were seen by the divers during the day throughout a nineday neap period in early May 2004.
Several specimens were caught with unidentified species of larval fish in the guts, both during fish frenzies at the night lights and also during the day while the divers were at decompression hang off. However, none were observed actively catching prey.
Molecular sequences
16S mtDNA and 18S nDNA gene sequences are being developed for this species, as part of a phylogenetic study to be published elsewhere (Gershwin and van Oppen, unpublished).
Remarks
Notably, several of the specimens are smaller than the holotype, and thus, presumably less mature, yet have a more complex branching pattern of the velarial structures. Some authors believed that velarial canal complexity was dependent on maturity ( Bigelow 1909; Stiasny 1930); this seems to be partly true in most medusae I have studied. In general, in my experience, some species have a propensity to develop more canal complexity than others; in the species with such tendencies, the canals of the more mature individuals are more complex than those of the less mature individuals. However, this is the first time that I have seen this phenomenon in reverse.
The rhopalial horns are an interesting character, not considered by previous workers. They are characteristic of the “Irukandji species” (i.e., Carukia spp., Malo maxima and other undescribed Malo spp., “Darwin Carybdeid”, and “Morbakka”), but are curiously absent in Tamoya haplonema (which is otherwise similar to the Irukandji species in many respects) and present in Carybdea sivickisi (which is wholly unlike the Irukandji species in all respects). They are absent in all other Carybdeida and Chirodropida I have studied ( Gershwin, 2005). In M. maxima , the horns are less robustly developed than in the Darwin Carybdeid, in which they are large, thick, devilhorncurved structures. The function of these structures, if any, is unknown.

| WAM |
Western Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.