
Fraseroscyphus macrogonus ( Trebilcock, 1928 )
|
publication ID |
https://doi.org/10.11646/zootaxa.4168.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:4869692E-590E-4A7F-8632-C4CFDC7A5AC6 |
|
DOI |
https://doi.org/10.5281/zenodo.3510426 |
|
persistent identifier |
https://treatment.plazi.org/id/03CC514D-6534-FFE4-A28E-E09CFD36FC23 |
|
treatment provided by |
Plazi |
|
scientific name |
Fraseroscyphus macrogonus ( Trebilcock, 1928 ) |
| status |
comb. nov. |
Fraseroscyphus macrogonus ( Trebilcock, 1928) View in CoL comb. nov.
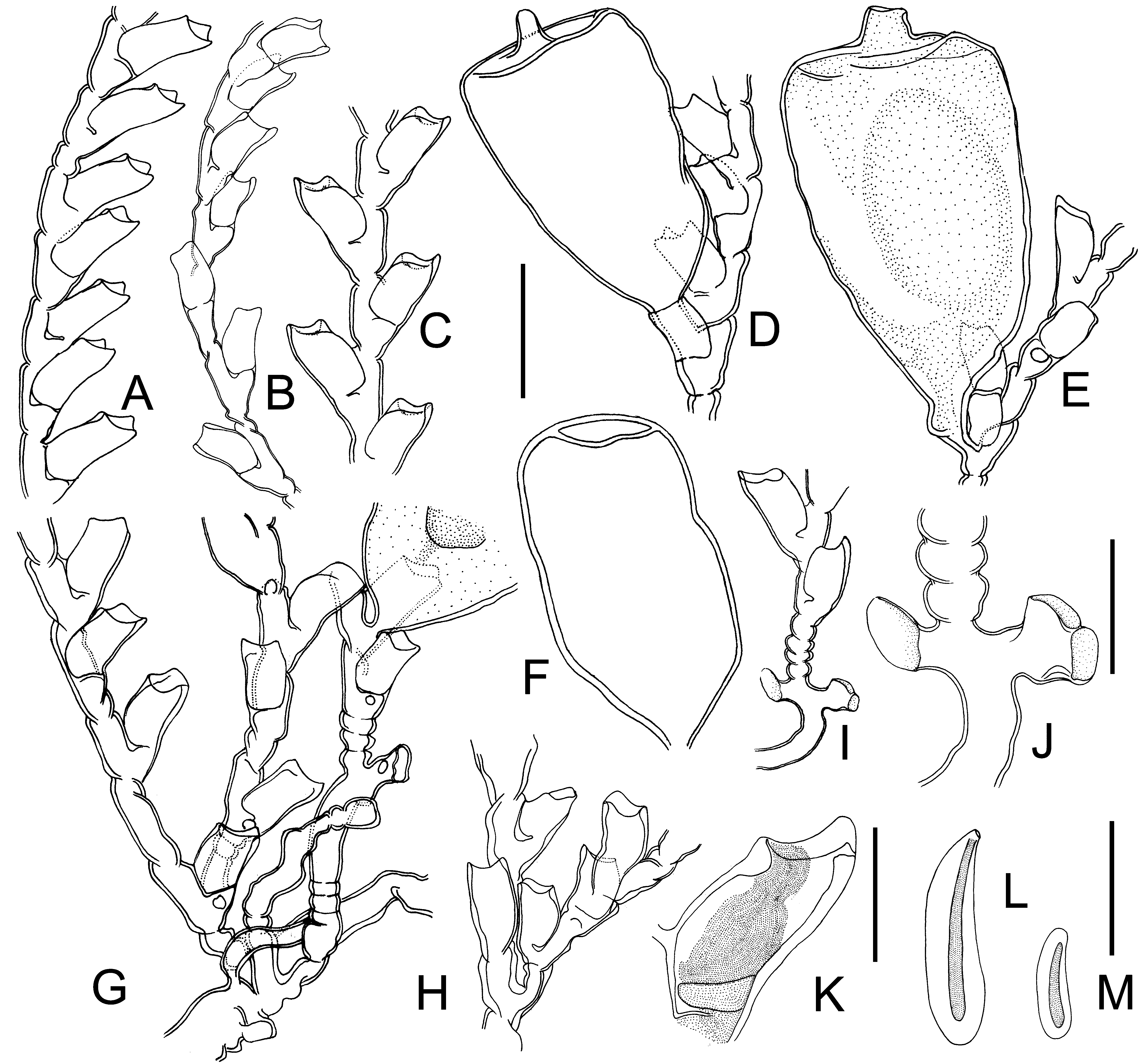
( Figures 3 View FIGURE 3 F–G, J, 6–7)
Sertularella macrogona Trebilcock, 1928: 11 View in CoL , pl. 1, figs. 4–4d.
Symplectoscyphus macrogonus View in CoL — Ralph, 1961a: 798, figs. 14a–b; 1961b: 109.— Dawson, 1992: 20.— Vervoort & Watson, 2003: 218, figs. 52A–D.
Sertularella macrocarpa — Stranks, 1993: 17.
? Symplectoscyphus macrogonus View in CoL — Millard, 1957: 219; 1964: 51; 1975: 316, figs. 102D–G; 1978: 199.— Gili et al., 1989: 104, fig. 29B.
? Symplectoscyphus sp.2— Galea & Schories, 2012: 56 View Cited Treatment , fig. 7J.
? Symplectoscyphus unilateralis — Galea et al, 2014, fig. 8A–D.
Type locality. St. Clair , Dunedin, New Zealand.
Type material. — NMV F57892 View Materials , syntype, two permanent slides ( NMV F57892.1 and.2), St. Clair , Dunedin, rock pools at low tide, 1923. IV– V, R. E. Trebilcock. Details : NMV F57892.1 ( Figure 3 View FIGURE 3 F), seven fragments, four male gonothecae (one originating from within the hydrothecal cavity, three growing below hydrothecae), two clinging organs, with 1–2 lobes, diameter 0.12–0.14 mm.—NMV F57892.2 ( Figure 3 View FIGURE 3 G), 14 fragments, five male gonothecae (two originating from within the hydrothecal cavity, two growing below hydrothecae, one detached).
Doubtful Chilean specimen. Re-examined based on original photomicrographs of Symplectoscyphus sp. 2 in Galea & Schories (2012): single fragment, alcohol sample fixed through formalin, with a broken gonotheca (possibly male) and a tendrils originating from the apertures of two sperate hydrothecae, Rivers Region , Corral, La Amistad (San Carlos), 39.85744° S, 73.44024° W, 5 m, 2010. V. 26, Horia Galea. GoogleMaps
Lectotype designation. —Lectotype, a fragment on the slide NMV F57892.1 as shown by an arrow in Figure 3 View FIGURE 3 F, the best fertile fragment, male.—Paralectotype, the remaining fragments on slide NMV F57892.1 and F57892.2, all fertile fragments are male.
Measurements. Full details are given in Table 6.
Description. Trophosome. Hydrocaulus small ( Figure 3 View FIGURE 3 F–G), monosiphonic, erect, slightly zigzag, with two to six transverse annulations at base, sparingly and irregularly branched ( Figure 6 View FIGURE 6 H), hydrocladium usually growing directly out of hydrocaulus beneath hydrotheca, without axillary hydrotheca ( Figure 6 View FIGURE 6 H), or infrequently originating from within the hydrothecal cavity. Regular and oblique nodes on hydrocaulus and hydrocladium, each internode slightly constricted at the end and bearing a hydrotheca; some hydrocauli or hydrocladia bending irregularly, resembling tendrils, hydrotheca absent or with infrequent hydrotheca, with abundant annulations ( Figure 6 View FIGURE 6 G). Hydrothecae alternately arranged in two longitudinal rows usually forming an angle of 40–60º ( Figure 6 View FIGURE 6 A–B), sometimes almost in one plane ( Figure 6 View FIGURE 6 C) or infrequently spirally ( Figure 6 View FIGURE 6 G); hydrotheca tubular, no wrinkles or lines on surface, a third of adcauline part adnate, bending outwards distally; some hydrothecae with fenestrae ( Figure 6 View FIGURE 6 E, G); margin with three cusps, one adcauline and two lateral abcauline, abcauline cusps larger; the inner perisarc of some cusps thickened ( Figure 6 View FIGURE 6 B–C), usually only an adcauline cusp and an outer abcauline cusp thickened ( Figure 6 View FIGURE 6 K); operculum composed of three triangular flaps forming a pyramid; retracted hydranth with poorly developed abcauline caecum ( Figure 6 View FIGURE 6 K).
Clinging organ. Only a pair of opposing clinging organs were found at the base of a hydrocaulus in the type material, resembling one or two bubbly lobes opening distally with a very thin perisarc ( Figure 6 View FIGURE 6 I–J), accompanied by annulations in nearby nodes.
Gonosome. Male gonothecae relatively large, four to five times the size of a hydrotheca, generally originating from within the hydrothecal cavity ( Figure 6 View FIGURE 6 D), or growing from the fenestra beneath a hydrotheca ( Figure 6 View FIGURE 6 E, G), pedicellate, with a short distal neck constricting towards the distal end; male gonothecae shaped like inverted cones, broadening upwards, top flattened ( Figure 6 View FIGURE 6 D). Female gonothecae absent in type material.
Nematocysts. Two types, capsule spindle-shaped, varying in length ( Figures 3 View FIGURE 3 K, 6L–M).
Distribution. Distributed on the east coast of South New Zealand: St. Clair Dunedin, Kaikoura drift (P.M.R.), Taylors Mistake near Christchurch (G. Knox) and Little Papanui near N.E. end of beach ( Trebilcock 1928; Ralph 1961a; Vervoort & Watson 2003). The validity of the records from South Africa ( Millard 1957, 1964, 1975, 1978) could not be determined as material was not re-examined. The Chile records identified as Symplectoscyphus unilateralis by Galea & Schories (2012) and Galea et al. (2014) should be investigated by re-examination of type material of S. unilateralis .
Remarks. In the catalogue of Cnidaria types in the Museum of Victoria ( Strank 1993), the names Sertularella macrogona Trebilcock, 1928 and Sertularia macrocarpa Bale, 1884 were reported to be transposed ( Vervoort & Watson 2003). This was verified after re-examination of the type material. We can confirm that NMV F57892 View Materials (as two slides, NMV F57892.1 and NMV F57892.2) is the type material for Sertularella macrogona , while NMV F58805 View Materials and NMV F58806 View Materials are the type slides for Sertularia macrocarpa .
Millard (1957, 1964, 1975, 1978) reported Fraseroscyphus macrogonus (as Symplectoscyphus macrogonus ) from South Africa. Her samples developed into shrubby colonies generated by abundant tendrils originating from within the hydrothecal cavity. They were characterized by three intrathecal teeth sometimes present alternating with marginal cusps. Several tendrils existed in the type material although they were overlooked by Trebilcock (1928). It was clear that Millard (1975) had not mistaken the thickened part of the marginal cusps as internal teeth, because in her illustration (Figure 102 F), she drew both characters alternating in the same hydrotheca. However, Millard (1957) mentioned that internal teeth were not a constant character and varied in her samples, and she could not rely on this character to separate her samples into a different species from Sertularella macrogona .
Two thickenings of marginal cusps in hydrothecae were very often found in the type material of Fraseroscyphus macrogonus ( Figure 6 View FIGURE 6 B–C, K), during re-examination using a compound microscope. We are certain that the thickening of marginal cusps is different from the intrathecal teeth illustrated by Millard (1975) for the South African records. Ralph (1961a) accepted Millard's records, and adopted the characters of tendrils and internal teeth ( Millard 1957) to the descriptions of her New Zealand samples. Although Ralph (1961a) mentioned that her samples did not have internal teeth, she drew an illustration (Figure 14b) showing two thickenings of marginal cusps in a specific hydrotheca, which followed the same arrangement as that in the type material ( Figure 6 View FIGURE 6 K). When Vervoort & Watson (2003) re-examined Ralph's samples, they also found no internal teeth, but drew thickenings of marginal cusps in Figure 52A. We conclude that thickenings of marginal cusps in hydrothecae are common in the type material and other New Zealand records ( Ralph 1961a; Vervoort & Watson 2003). The reported presence of three clear intrathecal teeth in the South African records indicates the need for further examination before the material can be reliably assigned to F. macrogonus .
We are also not sure about the records by Gili et al. (1989) that were based on samples without gonotheca, collected from Namibia, West Africa at depths of 167–429 m. Some characteristics of their samples are similar to certain colonies or fragments of Fraseroscyphus macrogonus , e.g., hydrocauli unbranched, slightly zigzag, with irregular oblique nodes, hydrothecae strictly arranged in one plane; however, the hydrothecae in their samples appear to be more slender than of the type material, clear intrathecal teeth are present in the hydrothecae, and this bathymetric distribution appears to be much deeper than that of other known specimens of F. macrogonus . Without further examination of samples with gonotheca, we cannot confirm whether these records are just normal variations of F. macrogonus or belong to a separate species.
Galea et al. (2014) assigned Sertularella macrogona (revised into Fraseroscyphus macrogonus here), Sertularella pulchella Jäderholm, 1904 , and Sertularella rentoni Bartlett, 1907 as synonyms of Symplectoscyphus unilateralis (Lamouroux, 1824) based on the characteristic shape of their gonotheca as reported in published data. Such data, however, had been interpreted differently in previous studies. Bartlett (1907), Ralph (1961a) and Vervoort & Watson (2003) did not mention the thickening of marginal cusps or intrathecal teeth in the samples of Sertularella rentoni . Ralph (1961a) and Vervoort & Watson (2003) regarded Sertularella rentoni as distinct from F. macrogonus . Galea et al. (2014) mentioned that Billard (1909) examined the type material of Symplectoscyphus unilateralis and illustrated clear intrathecal teeth. Galea et al. (2014) also illustrated clear intrathecal teeth in the record of Symplectoscyphus unilateralis from Chile. Symplectoscyphus sp. 2, another Chilian specimen, from Galea and Schories (2012) was later also treated as S. unilateralis by Galea et al. (2014) because of these shared characters. The original description of Sertularella pulchella Jäderholm, 1904 is poor, but from the illustrations, the hydrothecal arrangement and gonotheca are similar to F. macrogonus . As a result, only based on the type material of F. macrogonus (male) we examined, we could not make a conclusion about its relationship with the remaining nominal species provided by Galea et al. (2014). Our redescription of the type material of F. macrogonus hopefully will help to resolve these issues in future studies.

| NMV |
Museum Victoria |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Hydroidolina |
|
Order |
|
|
Family |
|
|
Genus |
Fraseroscyphus macrogonus ( Trebilcock, 1928 )
| Song, Xikun, Xiao, Zefeng, Gravili, Cinzia, Ruthensteiner, Bernhard, Mackenzie, Melanie, Wang, Shaoqing, Chen, Jinjing, Yu, Nan & Wang, Jianjun 2016 |
Sertularella macrocarpa
| Stranks 1993: 17 |
Symplectoscyphus macrogonus
| Vervoort 2003: 218 |
| Dawson 1992: 20 |
| Ralph 1961: 798 |
Symplectoscyphus macrogonus
| Gili 1989: 104 |
| Millard 1957: 219 |
Sertularella macrogona
| Trebilcock 1928: 11 |