
Luzulaspis kinakikir Tanaka, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4985.3.8 |
|
publication LSID |
lsid:zoobank.org:pub:BB22ACD7-0061-4EF1-BF3F-DF9A5CF477D6 |
|
DOI |
https://doi.org/10.5281/zenodo.5074183 |
|
persistent identifier |
https://treatment.plazi.org/id/B6558787-DE3E-7054-F1D6-FD1197B9FB5B |
|
treatment provided by |
Plazi |
|
scientific name |
Luzulaspis kinakikir Tanaka |
| status |
sp. nov. |
Luzulaspis kinakikir Tanaka sp. nov.
Japanese common name: Ainu-hoso-katakaigaramushi
( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Material examined. Holotype: JAPAN, / Hokkaido / Kamikawa-gun, Pippu / Kita 2 sen / on Carex miyabei / 6. vi. 2020 / coll. D. Sasaki; adult female mounted singly on a slide ( ELKU) . Paratypes: same data as for holotype, 7 adult females mounted singly (4 EUMJ, 3 ELKU) .
Description of adult female (n = 8)
Live appearance. Body of adult female elongate oval, usually flat. Before oviposition, entire dorsum greenish yellow or almost infested glass-like color and without visible wax ( Fig. 2b–d View FIGURE 2 ). Ovisac-producing adult female not observed.
Slide-mounted adult female. Body elongate oval, 4.5 (4.3–5.0) mm long, 1.5 (1.3–1.8) mm wide, approximately 3 times longer than broad, sides parallel, margin with a distinct but shallow indentation at each stigmatic cleft; anal cleft approximately 1/9 (1/8–1/10) of body length.
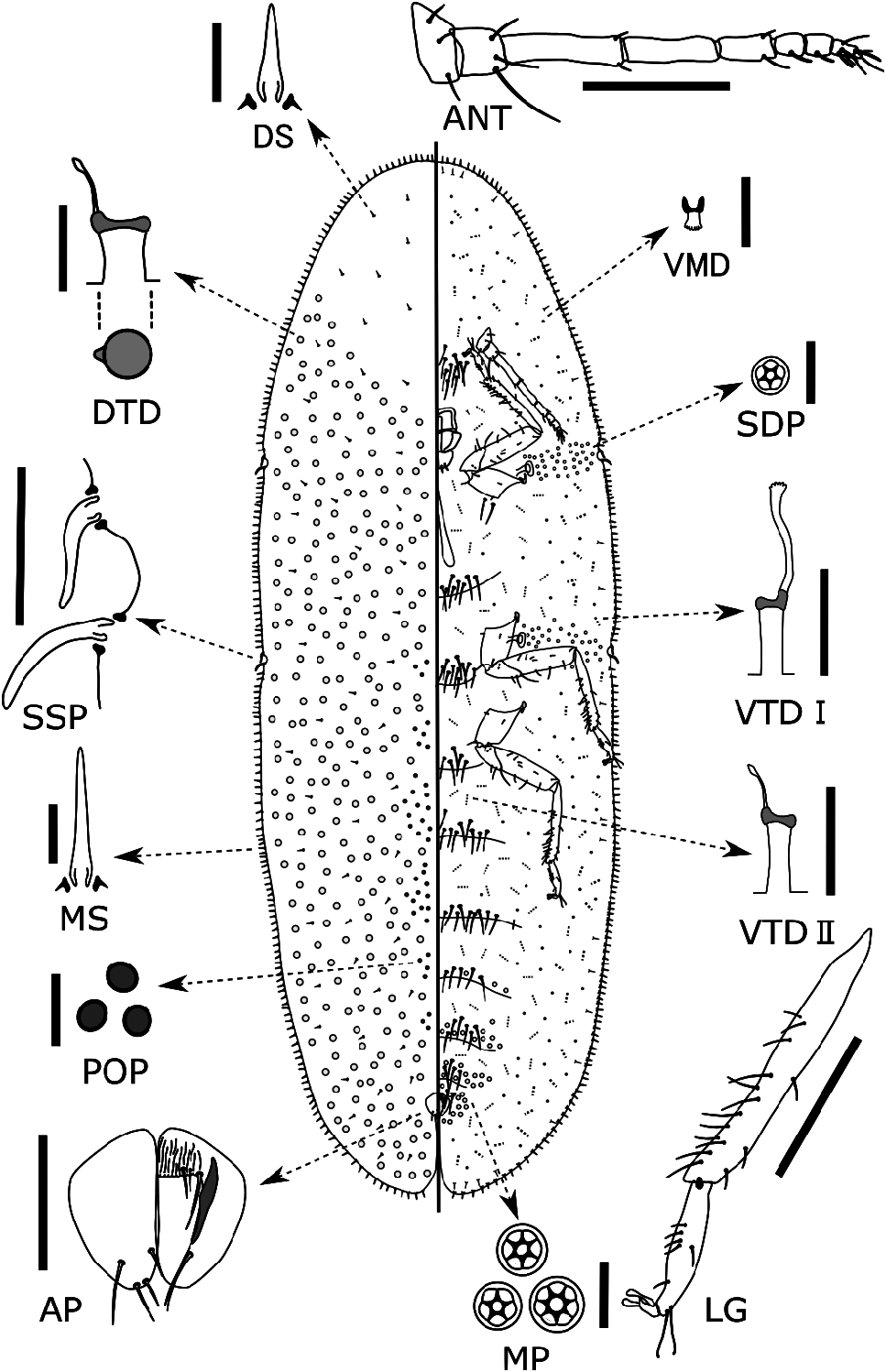
Dorsum. Derm membranous, dermal areolations absent. Setae spiniform, frequent, scattered throughout, each 7–10 (7–13) μm long with a well-developed basal socket; 7–9 (6–15) setae present within an area between antennal base and procoxa on the venter. Simple pores present, each 1–2 μm in diameter, distributed sparsely throughout. Preopercular pores oval to circular, each 3–4 (3–5) μm in diameter; present in a relatively wide band of approximately 100+ pores extending between anal plate and prothorax. Tubular ducts frequent throughout except for most of head ( Fig. 1 View FIGURE 1 ), each with a shallow but quite wide outer ductule [6–7 (5–9) μm wide and 4–6 (4–7) μm long] with a well-sclerotized invagination at inner end, and a thin inner ductule [<1 μm wide and 4–6 (3–10) μm long] with a small terminal gland, orifice obviously wider than those of type I ventral tubular ducts. Dorsal tubercles absent. Anal plates together quadrate but slightly rounded ( Fig. 1 View FIGURE 1 , AP; Fig. 3d View FIGURE 3 ), each plate 123–127 (120–138) μm long, 61–62 (57–76) μm wide, with a well-developed but small supporting bar, a slightly convex posterolateral margin and 4 apical or subapical setae. Ano-genital fold with 1 pair of well-developed setae, 0–2 small setae along anterior margin and 1 pair laterally. Anal ring bearing 6 setae.
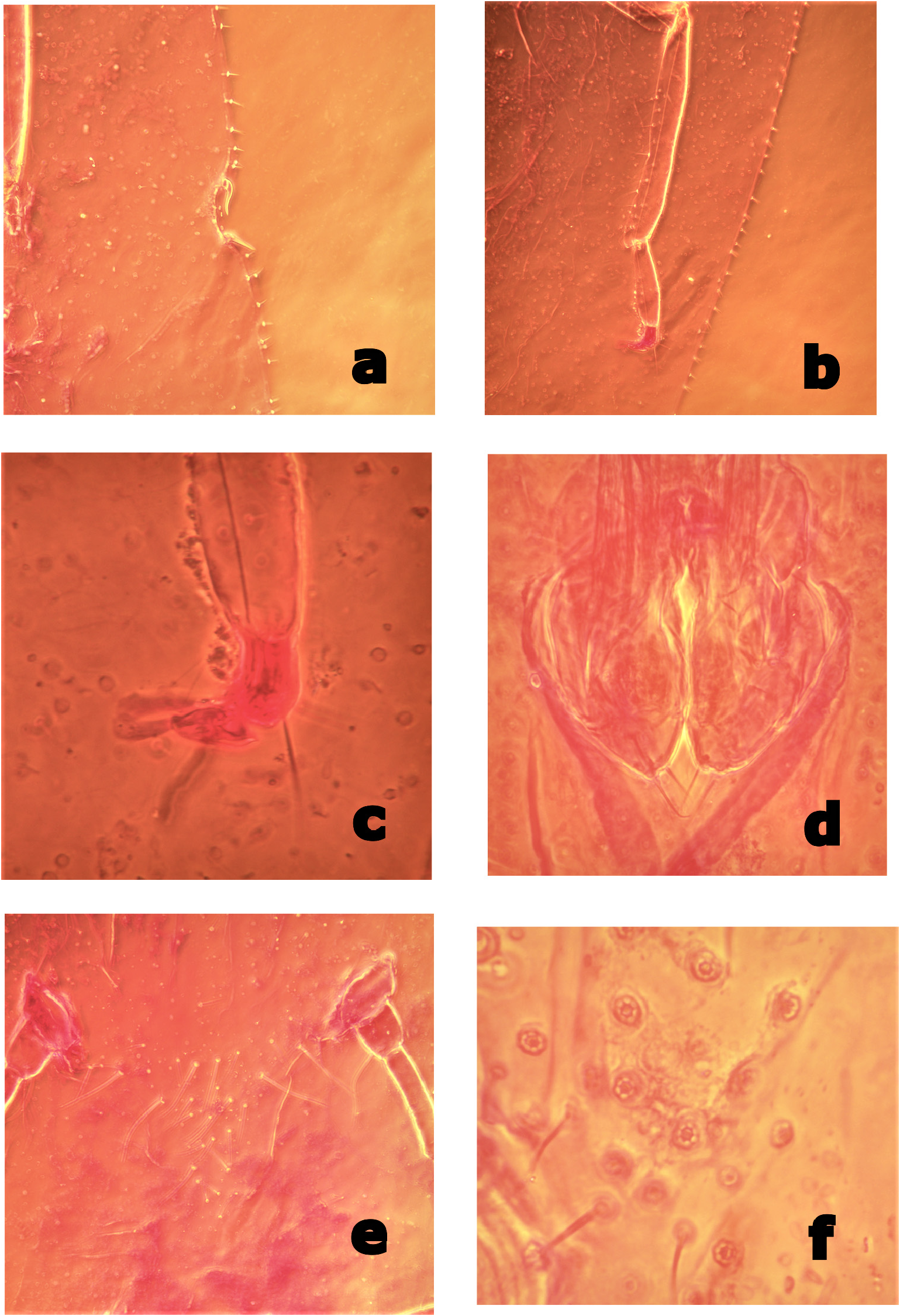
Margin. Marginal setae spiniform with well-developed basal sockets, mostly each 10–32 (9–35) μm long, notably shorter than stigmatic spines, normally in a single row; marginal setae on head and anal lobe slightly longer than those on lateral margin, each side with 30–33 (30–44) setae between anterior and posterior stigmatic clefts. Stigmatic clefts distinct, each containing 2 stigmatic spines; both spines 35–45 (32–55) μm long, each stout and bent, generally posteriorly but sometimes irregularly ( Fig. 3a View FIGURE 3 ), typically with a blunt apex. Eyespots not detected.
Venter. Derm membranous. Multilocular pores each 6–9 μm wide, with 5–7 (4–8), mostly 6 or 7 loculi ( Fig. 1 View FIGURE 1 , MP; Fig. 3f View FIGURE 3 ), present around genital opening and on medial areas of preceding 3‒5 abdominal segments. Spiracular pores each 4–5 (3–6) μm wide, mostly each with 5 loculi, present in a relatively broad band, 3–5 (3–8) pores wide, between margin and each spiracle; anterior bands each containing 17–22 (17–43) pores, posterior bands each with 28–33 (23–59) pores. Microducts scattered mainly on submarginal area but sometimes present on medial area of thoracic and abdominal segments. Tubular ducts of 2 types: type I each with outer ductule [3–4 μm wide and 6–10 (6–13) μm long], a slightly stout inner ductule [1–2 μm wide and 10–15 (10–20) μm long] and a flower-shaped terminal gland, present in outer submarginal area of head, thorax and abdomen; type II ducts each with outer ductule similar to type I [3–4 (2–4) μm wide and 5–8 (5–15) μm long], but with a short, filamentous inner ductule [<1 μm wide and 3–5 (3–8) μm long] and a minute terminal gland, present in medial and inner submarginal areas of head, thorax and abdomen. All thoracic and abdominal segments with a considerable number of long ventral setae in medial area, as follows: prothoracic segment, 18 (10–23); mesothoracic segment, 22 (9–23); metathoracic segments, 10 (6–13); abdominal segment I, 21 (9–21); II, 17 (9–17); III, 12 (8–15); IV, 12 (10–16); V, 7 (6–13); VI, 10 (5–10). With 28 (25–33) pairs of long interantennal setae between antennal bases ( Fig. 3e View FIGURE 3 ), 2–5 (2–7) pairs of long setae on posterior area of procoxae, 0 (0–6) pairs on anterior area of mesocoxae, and 0–2 (0–6) pairs on anterior area of metacoxae; relatively long submarginal setae [each 15–30 (9–30) μm] sparsely present along area inside margin; other setae short and fine, distributed over entire venter. Spiracles normal for the genus; peritreme widths: anterior spiracle 60–66 (52–67) μm, posterior spiracle 60–70 (60–78) μm. Legs well developed, each with a completely articulated tibio-tarsal joint ( Fig. 1 View FIGURE 1 , LG; Fig. 3 View FIGURE 3 , b) and an articulatory sclerosis; claw without a denticle; both claw digitules rather broad (Fig, 3, c) and slightly shorter than thin tarsal digitules. Hind trochanter + femur 437–444 (430–528) μm long, hind tibia 374–380 (350–438) μm long, and hind tarsus 158–172 (152–185) μm long; hind tibia with numerous relatively short setae (length subequal to tibia width). Antennae each 8-segmented, 606–632 (606–698) μm long. Labium 130 (113–134) μm wide, 60 (60–130) μm long.
Host-plants. Carex miyabei (Cyperaceae)
Remarks. The type specimens of Luzulaspis kinakikir sp. nov. have a large number (25 to 33) of well-developed long interantennal ventral setae; this contrasts with the morphological character state proposed by Hodgson (1994) as a diagnostic character of the genus (<30 well-developed long ventral setae between antennae), and differs from most of its congeners. However, most of the type specimens have fewer than 30 well-developed long ventral setae between the antennae, and only two of the paratypes have more than 30 (31 and 33 respectively).
Poaspis Koteja, 1978 is considered by some taxonomists to be the genus most closely related to Luzulaspis ( Çalýþkan et al. 2015; Hodgson 1994; Koteja 1979b). All the type specimens of L. kinakikir sp. nov. lack dorsal tubular ducts on most of the head ( Fig. 1 View FIGURE 1 ). This character state is similar to that in L. luzulae ( Dufour, 1864) , the type species of Luzulaspis , but different from that in the type species of Poaspis , P. jahandiezi ( Balachowsky 1932) . Therefore, we have tentatively placed L. kinakikir sp. nov. in Luzulaspis .
Koteja (1979b) split the members of Luzulaspis into five species groups, namely, Scotica, Luzulae, Bisetosa, Frontalis, and Grandis ( Çalýþkan et al. 2015). Luzulaspis kinakikir sp. nov. is considered to belong to the Scotica group because it has the following features: (i) strong, conical marginal setae in a single row; and (ii) dorsal setae small, conical, or nearly parallel-sided and subequal in length. Within this group, L. kinakikir sp. nov. is similar to L. filizae Kaydan, 2015 , in having: (i) setae on hind tibia relatively short (subequal or shorter than tibia width); (ii) head apex and anal lobe with marginal setae relatively longer than those on lateral margins; and (iii) stigmatic spines longer than other marginal setae. However, the new species differs from L. filizae as follows (character states of L. filizae are shown in parentheses): (i) multilocular pores each with 4–8, mostly 6 or 7, loculi (mostly each with 10–12 loculi); (ii) dorsal tubular ducts that are obviously wider than ventral tubular ducts (dorsal ducts same shape and size as the larger type of ventral ducts); and (iii) dorsal tubular ducts absent from head apex (sparsely present).
Biology. Luzulaspis kinakikir sp. nov. was found only on a few ramets of Carex miyabei in the bed of the Pippu River, where the same plants formed a dense cluster, and it is probably a rare species. The new scale species infests indented parts of the abaxial and proximal surfaces along two mid-lateral veins of young leaves, which are Mshaped in cross section ( Fig. 2a, b View FIGURE 2 ). While they were reared on the plants in the laboratory for a week, we observed L. kinakikir walking from old to new leaves of C. miyabei ( Fig. 2c, d View FIGURE 2 ), so the adult females are able to walk during the maturation stages before oviposition. The adult male and the life cycle of L. kinakikir are unknown.
Etymology. The specific epithet “kinakikir” is a new compound noun based on the language of the local indigenous Ainu people on Hokkaido Island. “ Kina ” refers to grass and sedge, and “Kikir” refers to a bug in the Ainu language. The epithet is used in apposition. In a few Ainu local dialects, “Kina-kikir” may also refer to the larvae of some lepidopteran species.

| EUMJ |
Ehime University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |