
Linderiella jebalae, Boix, Dani, Sala, Jordi, Escoriza, Daniel & Alonso, Miguel, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4138.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:8ABBDA5A-4F25-4990-BC3E-95E1CA1586AC |
|
DOI |
https://doi.org/10.5281/zenodo.6090743 |
|
persistent identifier |
https://treatment.plazi.org/id/03E4B919-7F25-6E58-8DE8-FB9B9F76F9A2 |
|
treatment provided by |
Plazi |
|
scientific name |
Linderiella jebalae |
| status |
sp. nov. |
Linderiella jebalae View in CoL sp. nov.
( Figs 1–12 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 )
Etymology. This species is derived from the word Jebala, corresponding to the mountainous region of Belad Jebala (“Le Pays Jbala”), in northwest Morocco, where the populations of this species are located. It is to be treated as a noun in the genitive.
Type locality. A temporary pond (BH1; 35°15’35”N, 05°25’28”W) near Moulay Abdessalam, province of Chefchaouen, region of Tanger –Tétouan–Al Hoceïma, Morocco. From this locality, several specimens were collected on 8 December 2011 by Daniel Escoriza and Dani Boix.
Type material. All material preserved in formaldehyde 4%.
Holotype. male; TL 7.9 mm, SL 7.2 mm; MNCN 20.04/10175.
Allotype. female; TL 10.1 mm, SL 9.3 mm; MNCN 20.04/10176.
Paratypes. 1) 1 male; MNCN 20.04/10177. 2) 1 male; MNCN 20.04/10178. 3) 1 male; MNCN 20.04/10179. 4) 1 female; MNCN 20.04/10180. 5) 1 female; MNCN 20.04/10181. 6) 1 female; MNCN 20.04/10182. 7) 3 males and 3 females; MNHN-IU-2014-7382. 8) 1 male; MZB 2015-8637. 9) 1 male; MZB 2015-8638. 10) 1 male; MZB 2015-8639. 11) 1 female; MZB 2015-8640. 12) 1 female; MZB 2015-8641. 13) 1 female; MZB 2015-8642.
Additional material examined. All material preserved in formaldehyde 4%, except when otherwise stated. 1) 10 males and 10 females; BH1; 0 8 December 2011; D. Escoriza & D. Boix coll.; LLAM-GB63. 2) 15 males and 15 females; BH2; 0 9 December 2011; D. Escoriza & D. Boix coll.; LLAM-GB65. 3) 11 males and 3 females fixed in ethanol 70%; WR1; 0 9 December 2011; D. Escoriza & D. Boix coll.; LLAM-GB66. 4) 18 males and 23 females; OT; 0 1 May 2012; M. Alonso coll.; MA. 5) 11 males and 17 females; AB; 0 4 May 2012; M. Alonso coll.; MA.
Diagnosis. Male. Two-segmented antenna with soft proximal segment and sclerotized distal segment; the latter longer than the former, slender and slightly curved with a row of transverse ridges on inner surface; proximal segment with a basiomedial lanceolate outgrowth ending in a long, narrow and acute distal process. Penial processes acute and narrow with slightly curved apex.
Female. Medial side of proximal segment of antenna bearing a basal, long filiform outgrowth ending in a warty apex. Genital segments smooth (without dorsolateral and lateral fields of setulae), and with highly developed cuticular papillae in the second segment. Brood pouch globose and ovate in ventral view and with a keel-shaped process directed posteriorly in its dorsal side.
Cyst. Spherical, covered with thick, blunt, sometimes truncated spines and with small pores.
Description. Adult male. General. Body of living specimens ivory or greenish white coloured, with some structures coloured in orange (mainly the medial border of thoracopods and genital and postgenital segments) ( Fig. 1 View FIGURE 1 ).
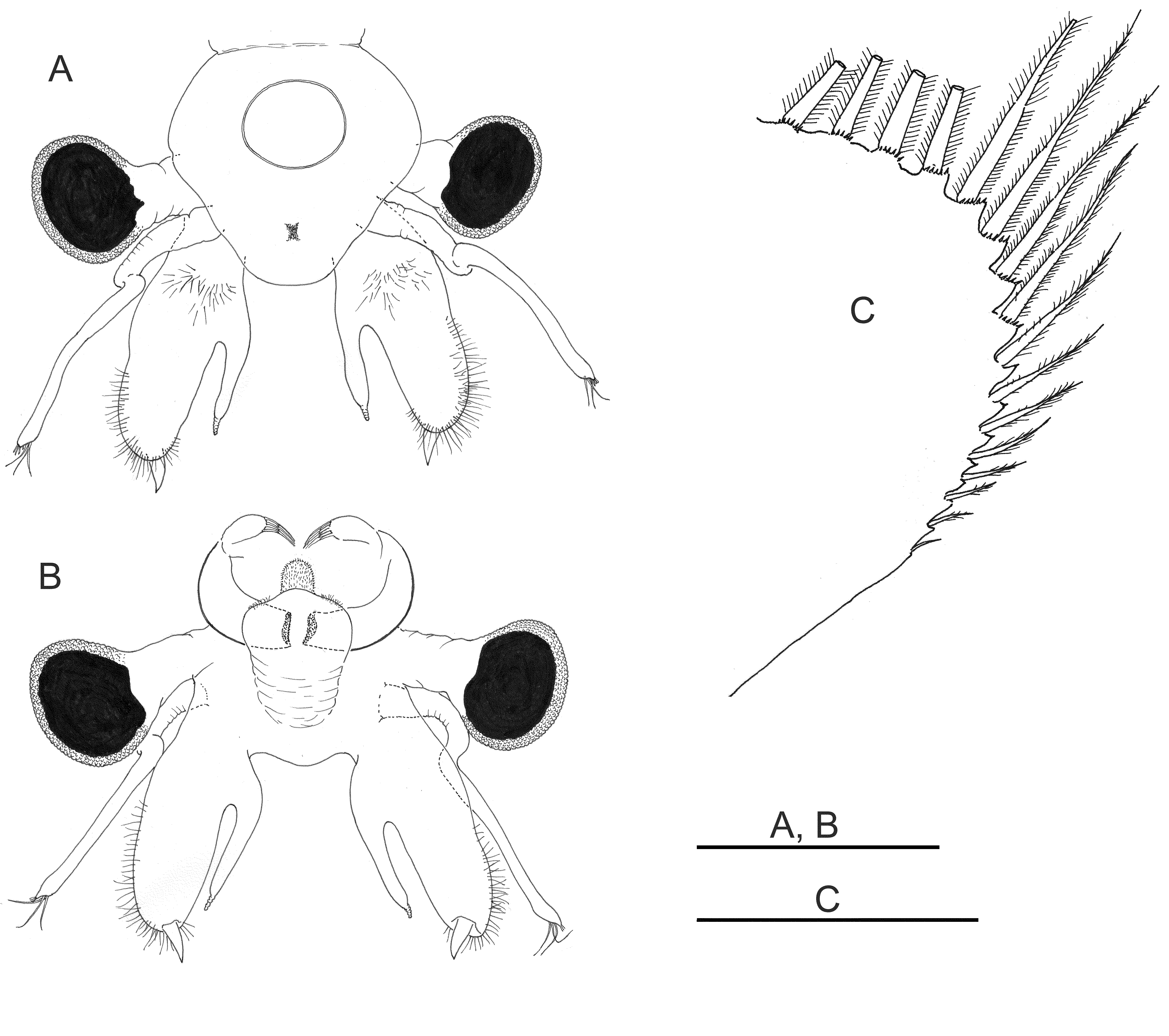
Head rounded with elliptic nuchal organ ( Fig. 2 View FIGURE 2 A).
Antennule about twice length of proximal segment of antenna, with three long subdistal setae and 12 aesthetascs ( Fig. 2 View FIGURE 2 F).
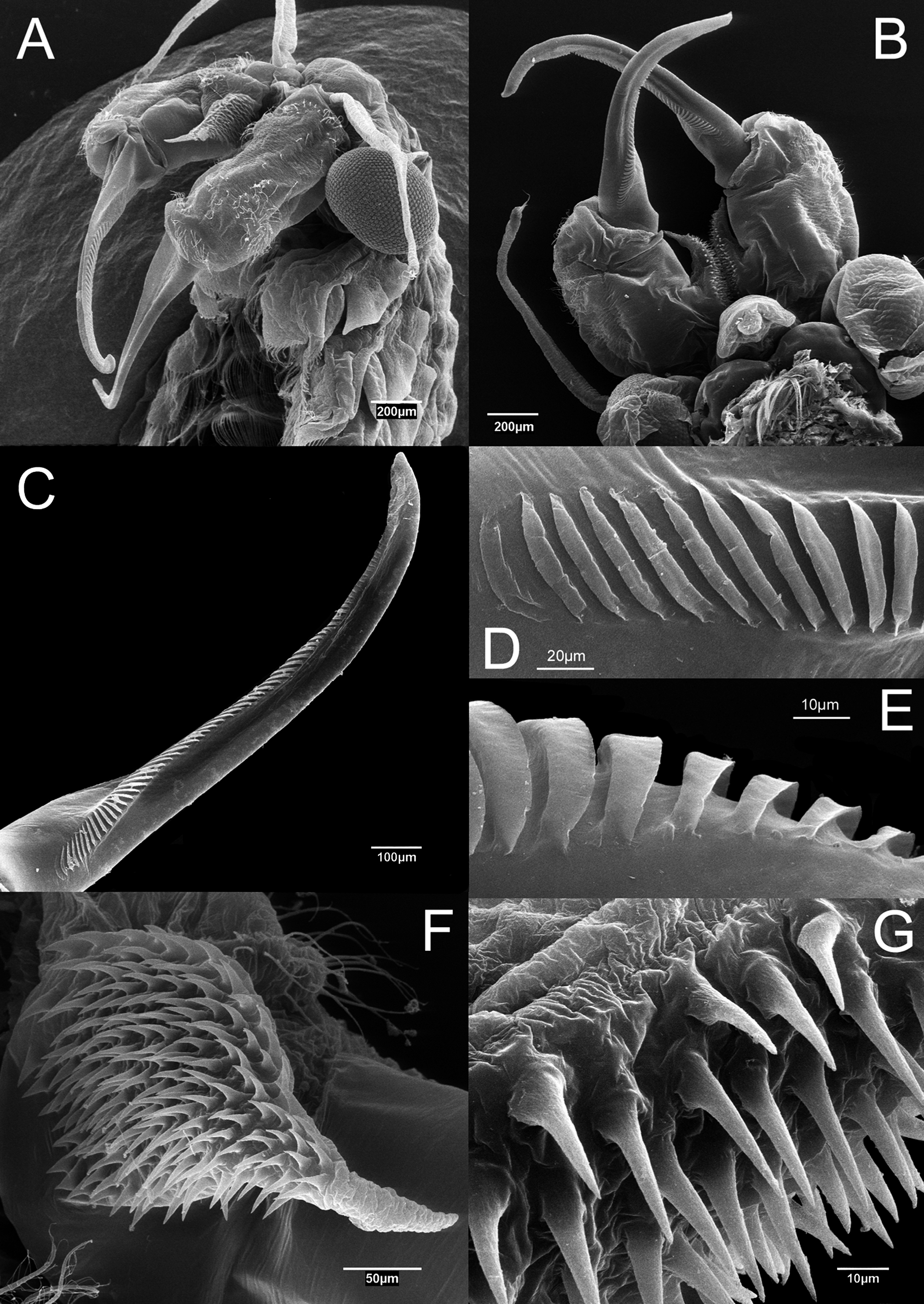
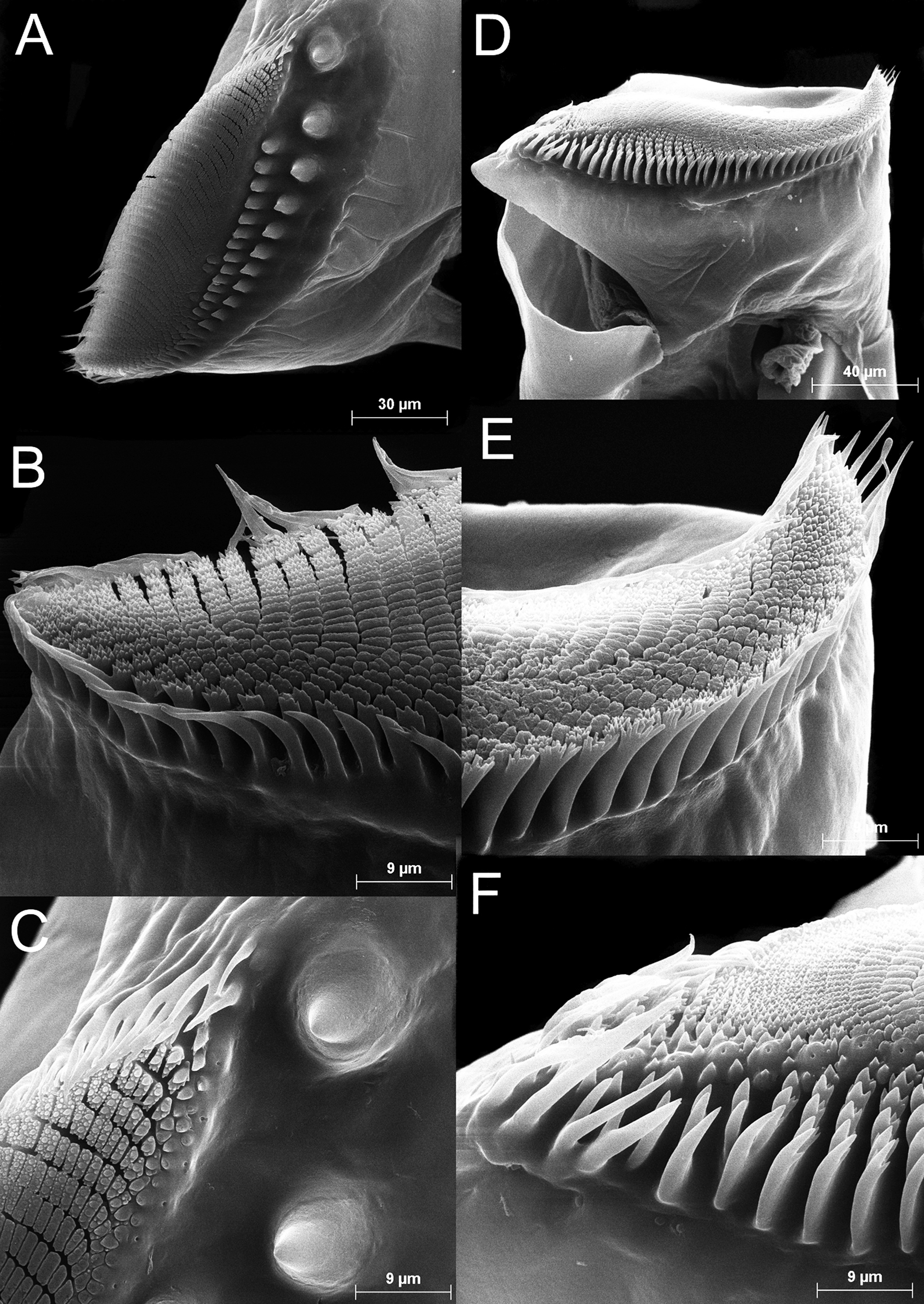
Antenna ( Figs 2 View FIGURE 2 A–B, 7A–E) 2-segmented, proximal segment soft, with patches of microtuberculate integument and sensillae distributed on dorsal and dorsolateral surface; basomedial outgrowth lanceolate in medial view, ending in long, narrow and acute process ( Figs 2 View FIGURE 2 C; 7F), and with medial side covered with smooth, slightly curved spinulae ( Figs 2 View FIGURE 2 C, 7F–G). Distal segment of antenna about 1.8 times longer than proximal segment, slender, narrowing distally, and slightly curved inward, with row of transverse ridges on medial surface ( Figs 2 View FIGURE 2 A–B; 7A– E); in its proximal part, row of transverse ridges bending towards ventral side of distal segment of antenna, continuing to base of distal segment for about a quarter of its length ( Fig. 7 View FIGURE 7 C).
Labrum ( Figs 2 View FIGURE 2 D–E, 7B) subquadrangular with setulose distal margins; short, fleshy setulose tongue-like process subdistally on dorsal (inner) side.
Mandibles as figured ( Fig. 8 View FIGURE 8 ), similar to those described for other species of the genus ( Mura 1996).
First maxillae each with 19–23 setae and posterior ventral spine ( Fig. 2 View FIGURE 2 G). Basal part of setae about 0.4 times length of distal part; posterior side of basal part of setae covered with strong spines loosely arranged in row. Posterior ventral spine small, devoid of setulae.
Second maxillae small, covered with fine setulae, with 2 or 3 distal setulose setae and small proximal spine medially directed.
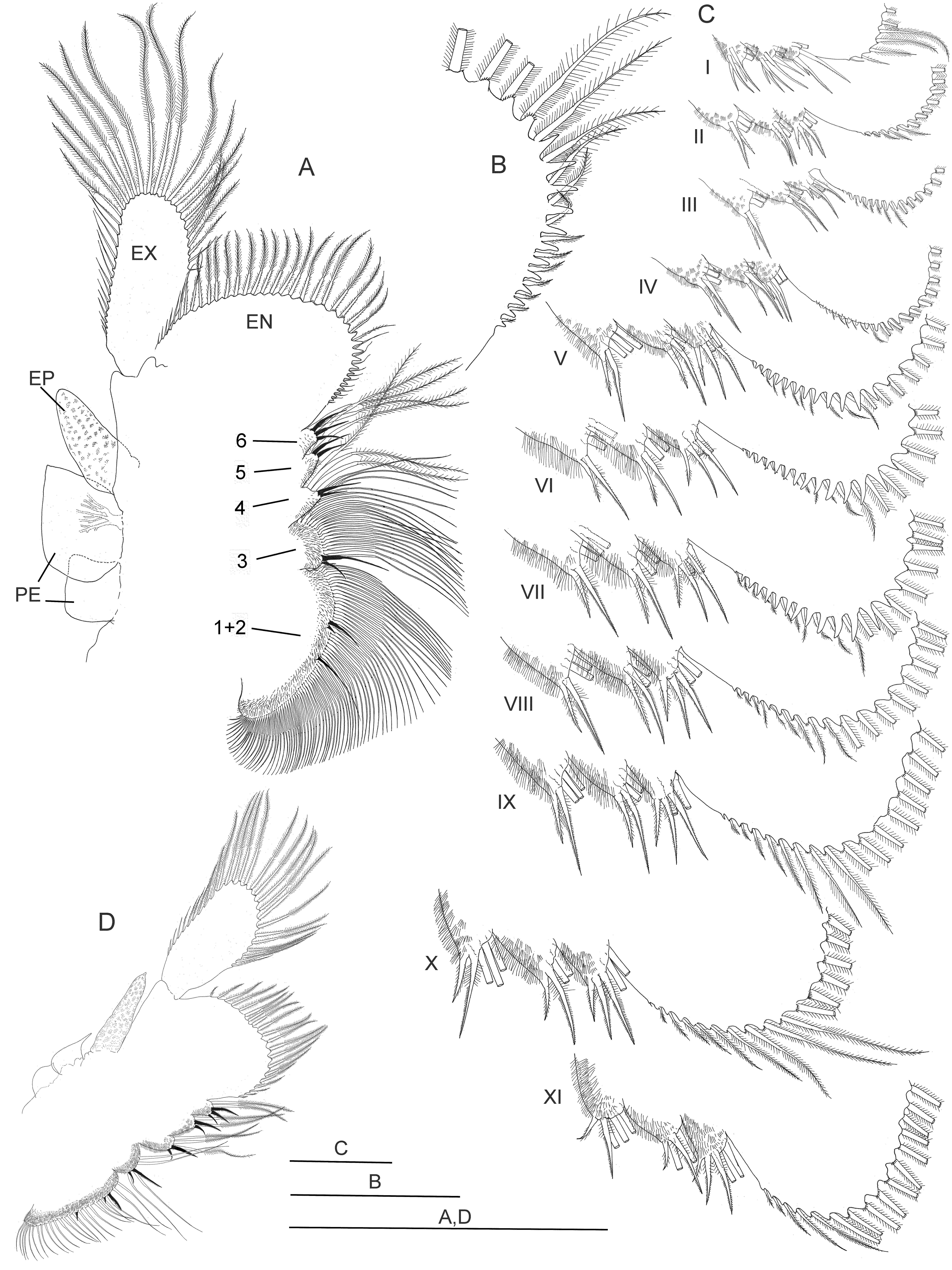
Thoracopods homonomous, with general structure typical for genus ( Fig. 3 View FIGURE 3 A, D). Thoracopods 5–7 the largest, being about 1.5 times larger than smallest thoracopods (T1 and T11).
Pre-epipodite (PE) subdivided into two leaf-like acuminate portions with smooth margins ( Fig. 3 View FIGURE 3 A). Length of distal portion about 1.5 times length of basal one. In T11, distal portion of pre-epipodite ending in a long acute extension ( Fig. 3 View FIGURE 3 D).
Epipodite (EP) digitiform, longer in relation to width from T1 to T10 (from about 2.3 to 2.6 times as long as wide); maximum width at base of epipodite; smooth margins and blunt end ( Fig. 3 View FIGURE 3 A). Epipodite of T11 more linear, about 4 times as long as wide, ending in mucronated tip ( Fig. 3 View FIGURE 3 D).
Exopodite (EX) oblong (slightly reniform in the first thoracopods), tending to be wider in relation to length from anterior to posterior thoracic limbs (from about 2.7 to about 2 times as long as wide) ( Fig. 3 View FIGURE 3 A, D). At T1, medial margin of exopodite almost naked (only its distal tenth has setae); on following thoracopods, medial margin becoming gradually covered with setae (from one-fifth at T2 to half its length at T11); rest of exopodite covered with setae. All setae plumose, except spine-like naked setae on proximal lateral margin. Pectinate scales at base of all setae, but especially developed in distal plumose setae.
Endopodite (EN) semi-oval, slightly expanded medially; endopodites with plumose setae ( Fig. 3 View FIGURE 3 A). Distinct acute projections at base of proximal plumose setae in T2–10 (sometimes also T11) ( Fig. 3 View FIGURE 3 B) reaching maximum size from T5–7. Pectinate scales at base of all setae, except when acute projections developed into acute spines (especially from T3–7) ( Fig. 3 View FIGURE 3 C).
Endite 1+2 of first 10 thoracic limbs with rounded edge covered with long filtering setae (lacking basal crown of pectinate scales) and with 3 anterior setae: the proximal one long and delicate, intermediate very short and spinelike, and the distal strong, spine-like and pectinate; the 2 distalmost anterior setae close to each other ( Fig. 3 View FIGURE 3 A). At T11, long filtering setae reduced in number and with strong basal scale posteriorly; proximal anterior seta very short and spine-like similar to intermediate one ( Fig. 3 View FIGURE 3 D).
Third endite ( Fig. 3 View FIGURE 3 A) about one-third of length of endite 1+2, with rounded edge covered with comb of long filtering setae, and with 2 unequal anterior setae close to proximal angle. At T11, endite 3 with typically 4 (3 or 4) filtering setae, and anterior setae situated in middle of endite ( Fig. 3 View FIGURE 3 D).
Fourth to sixth endites more or less conical in shape (more ovoid in T11). Long filtering setae on endites 4, 5 and 6 in the number of 3, 2 and 2 respectively. The number of anterior setae differs markedly on T1 from rest of thoracopods. On T1, endites 4, 5 and 6 with typically 4 (4–5), 6 (5–6), and 4 (4–5) anterior setae, respectively. On the rest of thoracopods, endite 4 and 5 with 2 anterior setae, and endite 6 with typically 3 (1–3) anterior setae ( Fig. 3 View FIGURE 3 A, C–D).
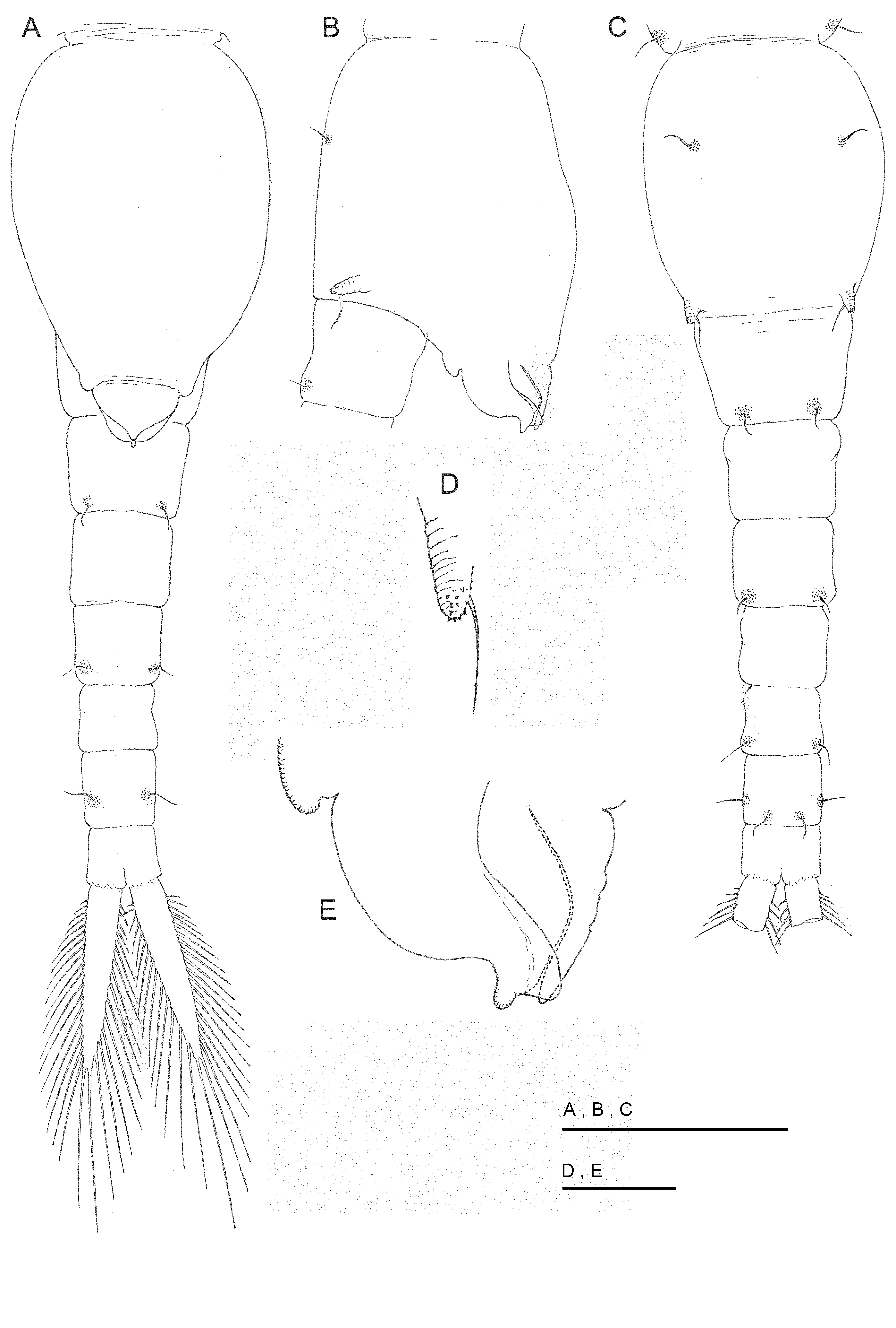
Genital somites ( Fig. 4 View FIGURE 4 A–B) swollen and partially fused; both somites with posterodorsal pair of cuticular papillae each provided with a sensilla, those of second genital somite slightly raised. Non-retractile portion of male gonopods elongated and cylindrical, extending to second postgenital somite ( Figs 4 View FIGURE 4 A–B; 9A–B), each bearing a medio-basal conical process with medial surface covered with short curved denticles ( Figs 4 View FIGURE 4 C, 9C–E). Eversible part of male gonopods half length of non-retractile portion, extending to third postgenital somite, conical with acute apex armed laterally with several spines ( Figs 4 View FIGURE 4 D; 9F–H).
Postgenital somites gradually decreasing in width. Second and fourth somites with pair of posteroventral cuticular papillae with sensillae ( Fig. 4 View FIGURE 4 A); first, third and fifth somites each with pair of posterodorsal cuticular papillae ( Fig. 4 View FIGURE 4 B); sixth somite with 3 pairs of cuticular papillae with sensillae, 1 posteroventral, 1 posterodorsal and 1 dorsolateral ( Figs 4 View FIGURE 4 A–B).
Cercopods ( Fig. 4 View FIGURE 4 A, E) more than 3 times length of telson. Setae at apex of cercopods implanted as figured ( Fig. 4 View FIGURE 4 E), longest as long as, or little longer, than cercopod itself. Surface of cercopods covered with tiny pectinate scales ( Fig. 4 View FIGURE 4 F).
Adult female. General. Body of living specimens dimmer than males, sometimes similarly coloured or rusty brown. Postgenital segments usually less coloured than males. Brood pouch transparent with 5 rusty brown dots (one ventral, two anterior dorsolateral, two posterior dorsolateral), oviduct dark blue ( Fig. 1 View FIGURE 1 ).
Head rounded, nuchal organ elliptic, as in male ( Fig. 5 View FIGURE 5 A).
Antennule 1.6 times longer than antenna ( Figs 5 View FIGURE 5 A–B; 10A, C).
Antenna subcylindrical, medial side of proximal segment of antenna bearing basal, distally directed filiform and acute outgrowth ending in warty apex (outgrowth 0.4 times length of antenna) ( Figs 5 View FIGURE 5 A–B, 10A–D); lateral margin of segment with several patches of microtuberculate integument, and with sparsely set sensillae distributed as figured. Distal segment of antenna reduced, conical and curved, tapering to a point.
Labrum ( Fig. 10 View FIGURE 10 E), mandibles, and first and second maxillae as in male.
Thoracopods similar to those of males. Acute projections at base of proximal marginal setae of endopodite absent or weakly developed (as in Fig. 5 View FIGURE 5 C).
Genital somites swollen and fused ( Figs 6 View FIGURE 6 B–C; 11A). Dorsolateral cuticular papillae present in both genital somites, those on second somite highly developed in digitiform process ( Figs 6 View FIGURE 6 B–D; 11D, F). Amplexial groove and dorsolateral and lateral sides of each genital somite devoid of hair-like setules ( Figs 6 View FIGURE 6 B–C; 11A, D–E).
Brood pouch (i.e., ovisac) globose and ovate in ventral view (sometimes cordiform when completely full of cysts), more or less fusiform in lateral view ( Fig. 6 View FIGURE 6 A–B). Brood pouch arising from both genital somites and extending to distal part of first postgenital somite (or reaching the proximal part of the second one). Dorsal side of brood pouch with keel-shaped process directed posteriorly ( Figs 6 View FIGURE 6 B, E, 11A–C). Pore opening subdistal, enclosed between two characteristic lips: ventral one ending in acute process, whereas dorsal one ending in more or less trilobed shape, with acute central process curled dorsally, and 2 blunt lateral processes directed caudally ( Figs 6 View FIGURE 6 A– B, E, 11A–C).
Cercopods less than 3 times length of telson ( Figs 6 View FIGURE 6 A; 10F). Surface of cercopods covered with tiny pectinate scales (as in Fig. 4 View FIGURE 4 F). Disposition and length of setae of cercopods as in males.
Cyst. Spherical; egg shell covered with spines 24 µm long (range = 19–38 µm, n = 46). Spines thick, blunt, sometimes truncated, covered with small pores ( Fig. 12 View FIGURE 12 ). Average cyst diameter, including spines, 300 µm (range = 275–313 µm, n = 46). Average spine density of 20 spines per 0.01 mm 2 (range = 14–28 spines per 0.01 mm 2, n = 5).
Size. Small-sized fairy shrimps. Males: TL, 6.8–9.1 mm (mean = 8.0 ± 0.5 mm, n = 23). Females: TL, 8.7–10.6 mm (mean = 9.5 ± 0.4 mm, n = 23). Mature females significantly longer (Student’s t-test, t 44 = -10.04; P <0.00001) than mature males.
Differential diagnosis. Males of Linderiella jebalae sp. nov. are not easily distinguished from males of other species of the genus. The antennal distal segment is falciform as in L. africana , L. massaliensis and L. baetica (contrasting with that of L. occidentalis and L. santarosae , which is more or less straight), but its maximum curvature is located at the distal third ( Figs 2 View FIGURE 2 A–B, 7A–C), whereas in the other species, the maximum curvature is in the middle or in the proximal third. Moreover, the row of transverse ridges of this antennal segment has a sinuous shape, being the proximal section, before the bend, one quarter longer than the distal section, after the bend ( Fig. 7 View FIGURE 7 C). All other species of the genus present a bend in the row of transverse ridges, but they lack the proximal straight section before it, with the exception of L. africana , in which some individuals have a very short section, less than one fifth of total length. The morphology of the basomedial outgrowth of the antennae also allows to separate the males of L. jebalae sp. nov. from the other species. The presence of a long distal process in the antennal outgrowth distinguishes the new species from L. occidentalis , L. baetica and L. africana , which do not present any process. L. santarosae and L. massaliensis can present a distal process in the antennal outgrowth, but its morphology is different from that of L. jebalae sp. nov., which is longer and thicker than in the other two species. In L. santarosae the process is short and curved ( Thiéry & Fugate 1994), whereas in L. massaliensis , only some individuals have a short distal process ( Thiéry & Champeau 1988). In L. jebalae sp. nov., all the individuals observed from different populations presented this distal process. Finally, the posterodorsal cuticular papillae of the second genital segment are slightly raised, while in the others species of the genus they are not significantly different from the rest of cuticular papillae of the postgenital segments.
In contrast, females of L. jebalae sp. nov. show several morphological features that clearly distinguish them from the other species of the genus. The long, filiform and acute basal outgrowth of the antenna, ending in a warty apex, clearly distinguish this species from the others; only L. massaliensis has a long acute outgrowth, but it is shorter than that of L. jebalae sp. nov. The antennal outgrowth of the latter species is about 3 times longer than distal segment of the antenna, while in L. massaliensis this proportion is about 1.4. Lateral and dorsolateral setulae of amplexial groove and genital segments are absent in the new species, in contrast with the other Palaearctic species of the genus. Moreover, the cuticular papillae on the second genital segment are clearly more developed than in the other species of the genus. The shape of brood pouch and its insertion in the genital segments also allow separation of this new species; it is ovoid in ventral view, and arises at the beginning of the first genital segment, not at the end of it, as in the other Palaearctic Linderiella . The presence of a keel-shaped process directed posteriorly in dorsal side of brood pouch is also only present in L. jebalae sp. nov.
The ornamentation of thick, blunt, sometimes truncated spines covered with small pores differentiates the cysts of L. jebalae sp. nov. from the cysts of the other species of the genus, which present a wide spectrum of spine morphologies (acute, tulip-shaped or trumpet-shaped spines).
Distribution and habitat. The species occurs in a mountainous region of the western Rif mountains between 719 and 1006 m above sea level ( Fig. 13 View FIGURE 13 ). The climate of the region is Mediterranean type (Csa in Köppen-Geiger classification; Peel et al. 2007) with an average annual temperature between 14.5–16.6° C and an average annual precipitation of 906–1052 mm. This climate is classified as humid (aridity index ranging between 0.67 and 0.89) based on UNEP (1997). The species inhabits temporary ponds in areas of cleared forest, mainly composed by cork oaks ( Quercus suber L.).
Ponds are small sized, with an average area of 81.1 m 2 (range = 11.4–174.2 m 2; n = 3) and shallow (mean depth = 29.9 cm; range = 6.5–51.1 cm; n = 3). Water can be clear or turbid by suspended clay in different ponds. Water mineralization is low–moderate (mean water electrical conductivity = 191.4 µS cm -1; range = 35.5–609 µS cm -1; n = 4). During daytime (11–16 h) between December and February, the mean water temperature was 12.7°C (range = 8.0–16.4°C; n = 4), mean pH was 6.8 (range = 6.0–7.7; n = 4) and mean dissolved oxygen saturation was 137.9% (range = 103.9–157.4%; n = 4). Clear water ponds contain well developed submerged macrophyte community, which explains the high values of dissolved oxygen. Accompanying aquatic fauna from several ponds is summarized in Tab. 1.
IUCN Red List status. Although this species appears in temporary aquatic habitats (and hence, some populations in diapause could have been overlooked), the survey of the area identified only five populations. Thus, Linderiella jebalae sp. nov. meets the IUCN Red List criteria ( IUCN 2001) for considering the species as Vulnerable, with an area of occupancy less than 2000 km 2, less than 10 locations, and extreme fluctuations in the area of occupancy (VU B2ac(ii)). The fact that L. jebalae sp. nov. has been found exclusively in temporary ponds makes them susceptible to extinction due to the existing threats to these habitats ( Grillas et al. 2004).
Ph. PLATYHELMINTHES
Cl. Turbellaria Typhloplanidae indet. Ph. ANNELIDA
Cl. Oligochaeta Tubificidae (Naidinae) indet. Ph. ARTHROPODA
Cl. Branchiopoda Alona affinis
Cl. Copepoda Eudiaptomus cf. chappuisi
Diaptomus sp.
Acanthocyclops sp.
Cl. Ostracoda Candona neglecta
......continued on the next page Eucypris virens Paralimnocythere psammophila
Cl. Arachnida Eylais sp.
Cl. Insecta
O. Ephemeroptera Cloeon gr. dipterum O. Hemiptera Microvelia pygmea Hesperocorixa moesta Sigara lateralis Sigara nigrolineata nigrolineata Parasigara infuscata Parasigara transversa Corixa iberica Notonecta maculata Notonecta meridionalis O. Coleoptera Gyrinus dejeani Dytiscus circumflexus Ilybius sp.
Agabus nebulosus Porhydrus vicinus Hydroporus pubescens Hydroporus planus Hydroporus obsoletus Graptodytes flavipes Berosus affinis Dryops striatellus O. Trichoptera Limnephilidae indet. O. Diptera Ochlerotatus rusticus Culiseta fumipennis Zalutschia sp.
Ph. CHORDATA
Cl. Amphibia Salamandra algira Hyla meridionalis Pelophylax saharicus
Cl. Reptilia Emys orbicularis







| MNCN |
Museo Nacional de Ciencias Naturales |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |