
Colocasiomyia leucocasiae Toda, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5079.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:2B118B81-7353-4A2C-A892-5466DFC83230 |
|
DOI |
https://doi.org/10.5281/zenodo.5786197 |
|
persistent identifier |
https://treatment.plazi.org/id/03D75C2D-4546-2E0D-FF52-62D3B057B60D |
|
treatment provided by |
Plazi |
|
scientific name |
Colocasiomyia leucocasiae Toda |
| status |
sp. nov. |
15) Colocasiomyia leucocasiae Toda View in CoL , sp. nov.
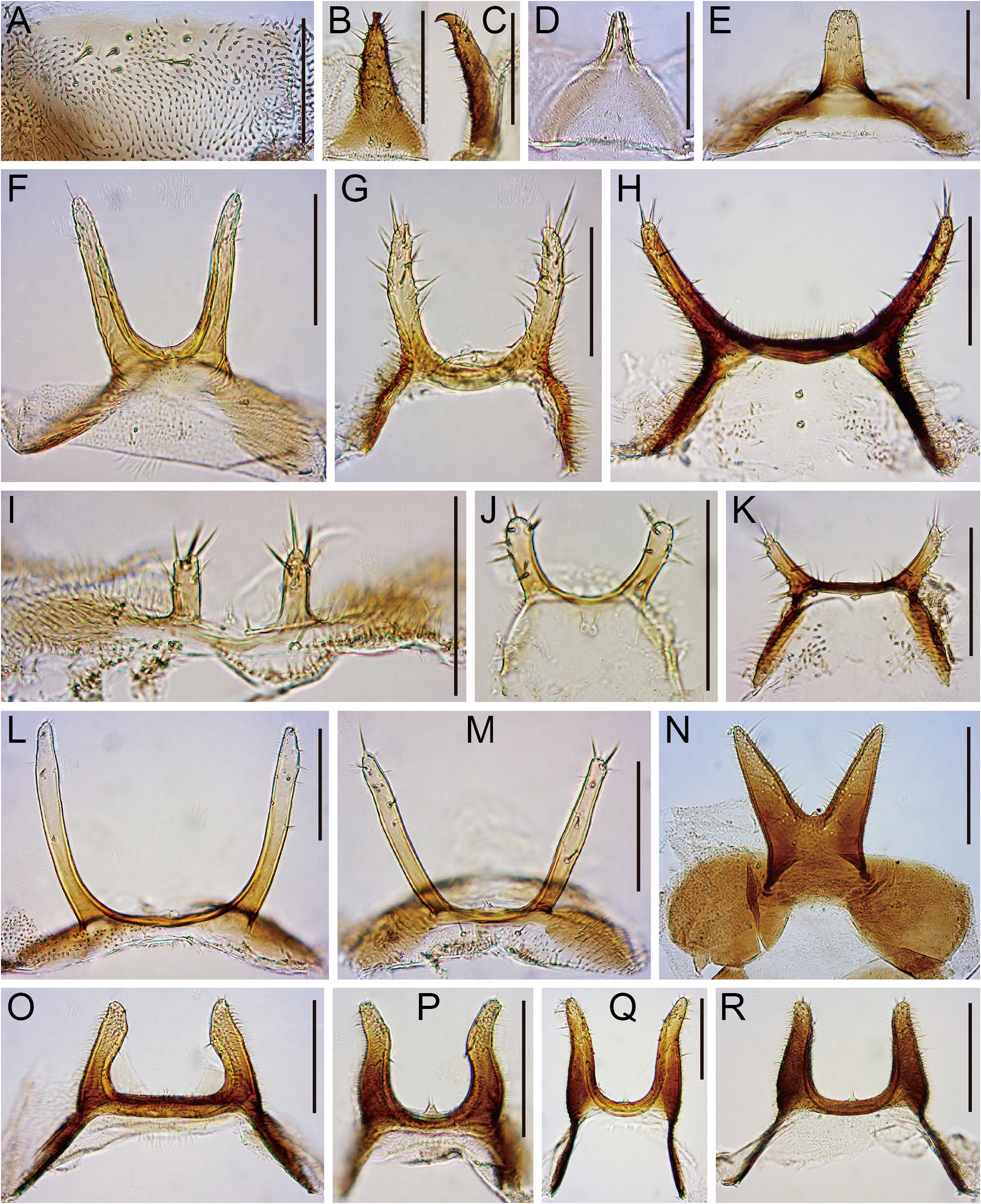
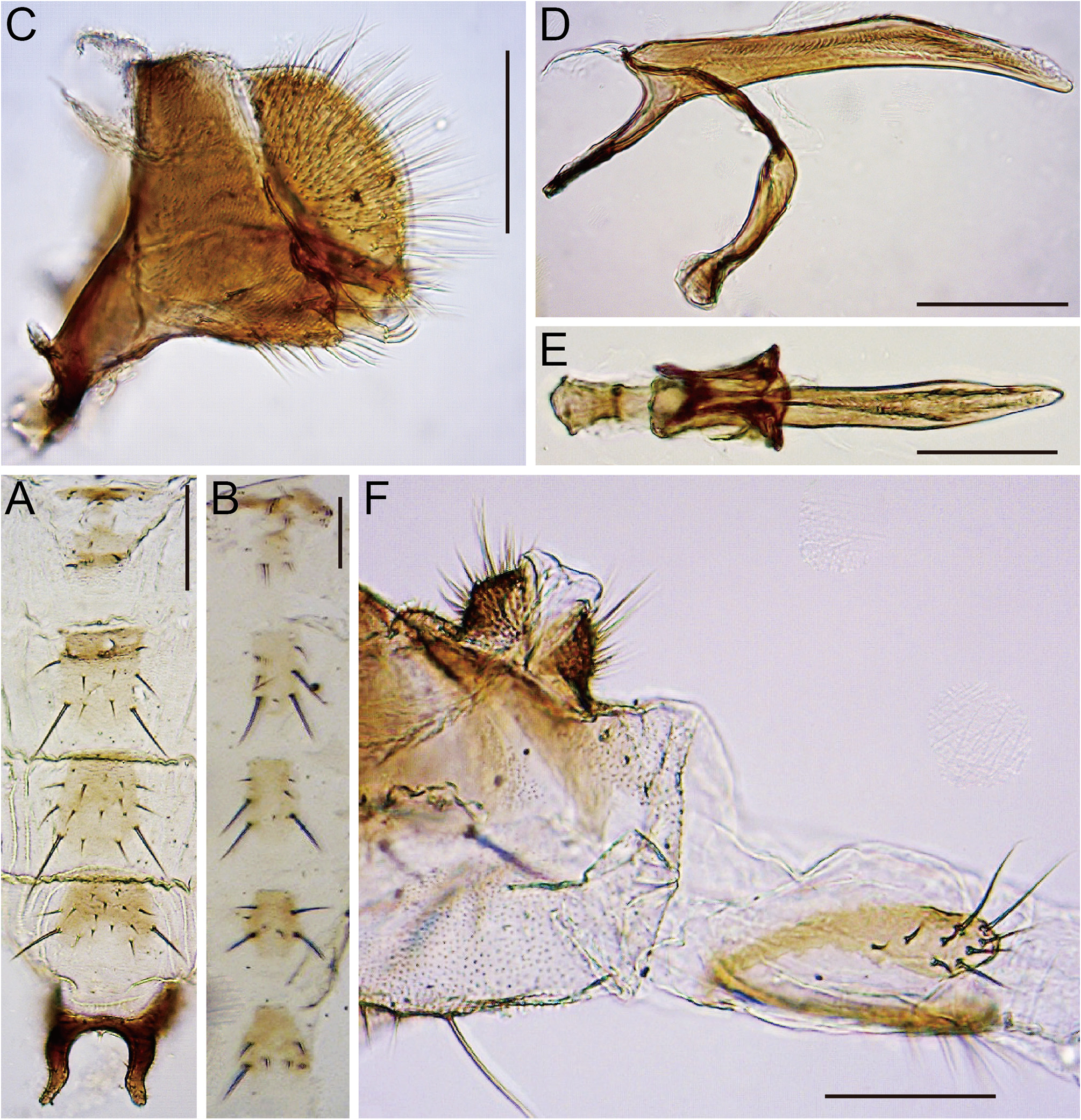
( Figs 9R View FIGURE 9 , 15O View FIGURE 15 , 29 View FIGURE 29 )
Colocasiomyia sp.1 aff. diconica: Sultana et al., 2006: 694 ; Takano et al., 2011: 22; Fartyal et al., 2013, Fig. 6 View FIGURE 6 .
Diagnosis. Apically blunt, heavy, peg-like costal setae in middle row interspersed with weak, trichoid ones ( Fig. 29A View FIGURE 29 ). A pair of processes on male abdominal sternite VI nearly straight and parallel ( Fig. 9R View FIGURE 9 ). Surstylus as broad as ventro-apical lobe of cercus, with 5–6 recurved teeth apically and 1–2 minute setae subapically ( Fig. 29D View FIGURE 29 ). Epiproct and hypoproct large; hypoproct thicker than width of oviscapt in lateral view ( Fig. 29G View FIGURE 29 ).
Description (♂ and ♀; not repeating characters common to C. xenalocasiae ). Head. Supracervical setae 5–6 per side. Cibarial, posterior sensillum 1 per side. Supralateral seta outside prementum 1 per side.
Legs. Mid tibia with approximately 2 apical, stout setae.
Abdomen. Male abdominal sternites III and IV wider than long, medially notched on posterior margin ( Fig. 29B View FIGURE 29 ). Female abdominal sternite VI wider than long, medially notched on posterior margin ( Fig. 29C View FIGURE 29 ).
Male terminalia. Epandrium pubescent except for anterior and ventral margins and anteroventral elongation, posteroventrally less extended, with 1–3 setae on lateral to dorsal portion and 16–19 setae on ventral portion of each side ( Fig. 29D View FIGURE 29 ). Cercus pubescent except for anterior margin and ventral 1/5, with 37–41 setae ( Fig. 29D View FIGURE 29 ). Oviscapt about 4 times as long as wide, with 12–13 ovisensilla ( Fig. 29G View FIGURE 29 ).
Indices (range of 10♂ and 10♀ paratypes): FW/HW = 0.50–0.71, ch/o = 0.31–0.67, prorb = 0.93–1.16, rcorb = 0.18–0.32, vb = 0.27–0.54, dcl = 0.46–0.70, sctl = 0.63–0.83, sterno = 0.34–0.63, orbito = 0.37–0.74, dcp = 0.79–1.23, sctlp = 0.80–1.02, C = 1.71–2.46, 4c = 1.14–1.53, 4v = 1.79–2.47, 5x = 1.31–2.13, ac = 2.68–3.51, M = 0.40–0.74.
Puparium (3rd instar larva). Segments with stout spicules on ventral surface; anterior spiracle sessile, with a bundle of short branches; caudal segments elongate, with small spicules, ending in a pair of posterior spiracles ( Fig. 29H–J View FIGURE 29 ). Mouth hook triangularly expanded medioventrally in lateral view; distal blade shorter than basal portion, apically more or less roundish, strongly curved downward, with two lines of blunt teeth on submedial portion of ventral margin ( Fig. 29K View FIGURE 29 ).
Holotype. ♂ ( MZB), “ Bogor Botanical Garden , Bogor, West Java, Indonesia, 9.i.2004, ex Leucocasia gigantea, K.T. Takano ”.
Paratypes. Indonesia: 11♂, 11♀, same data as the holotype ; 10♂, 10♀, Madakaripura waterfall, Probolinggo , East Java, 29.xii.2003, ex L. gigantea, K.T.Takano leg .; 10♂, 10♀, Kerinci National Park , Jambi, Sumatra, 7.xii.2004, ex L. gigantea, K.T. Takano leg. ( MZB, SEHU) . Malaysia: 1♂, Gua Ikan, Dabong, Kuala Krai , Kelantan, 5°21'15"N 102°01'37"E, 15.iii.2016, ex L. gigantea, S.Y. Wong leg GoogleMaps .; 10♂, 10♀, ditto except 17.iii.2016 ( SEHU) GoogleMaps .
Another specimen examined. Indonesia: 1♂, Sungai Lubuk Paraku , West Sumatra, 970 m a.s.l., 4.i.2004, ex Colocasia esculenta, K.T. Takano leg. ( SEHU) .
Distribution. Peninsular Malaysia, Sumatra (West Sumatra), Java (West and East Java).
Remarks. This species morphologically resembles C. diconica in having the epandrium less extended posteroventrally ( Figs 27C View FIGURE 27 , 29D View FIGURE 29 ) and the short oviscapt about 4 times as long wide ( Figs 27F View FIGURE 27 , 29G View FIGURE 29 ), but can be distinguished from it by the diagnostic characters.
Etymology. Referring to the main host plant.
Biogeography, flower-breeding ecology and evolution
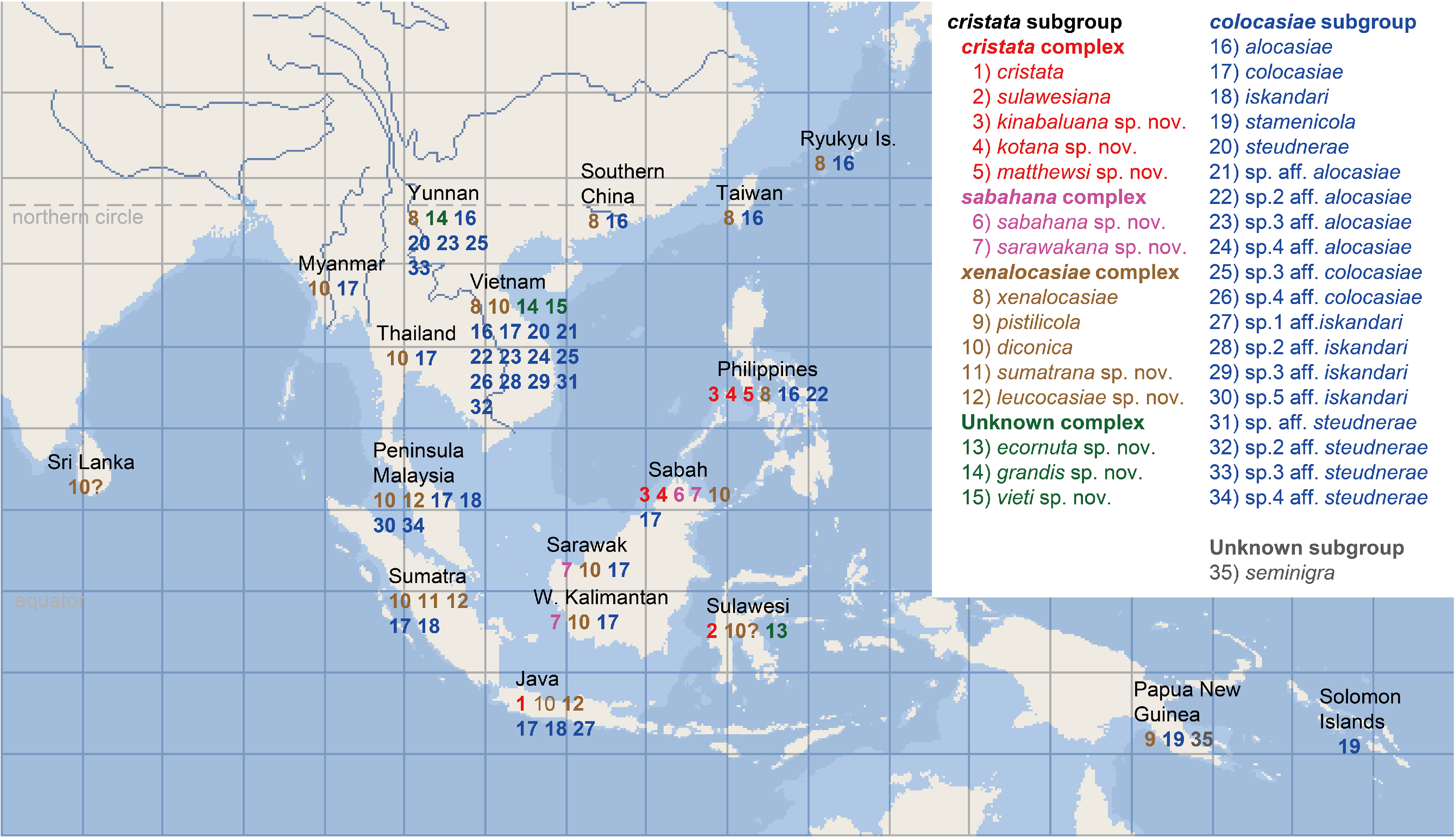
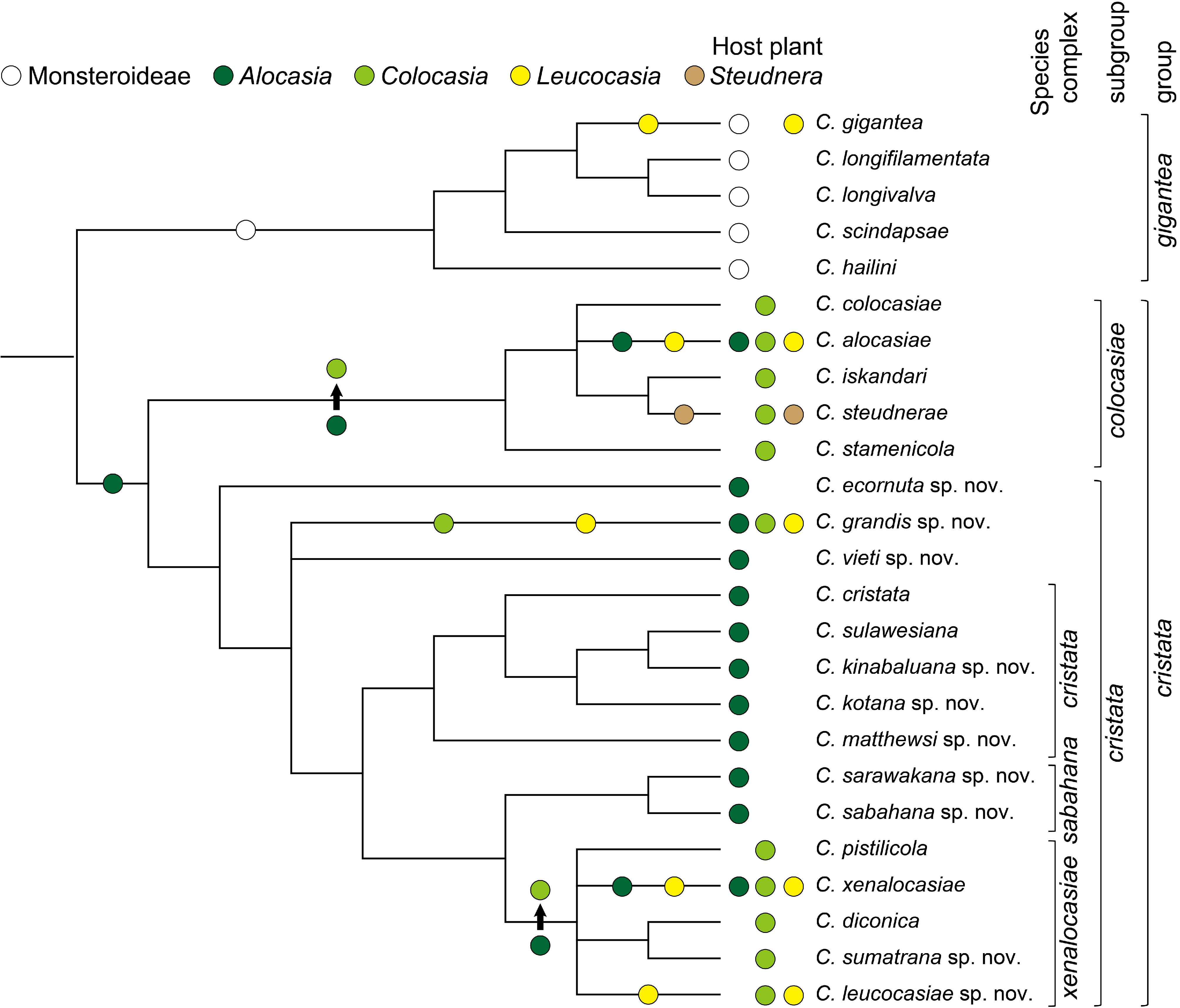
Based on up-to-date collection records of Colocasiomyia species , the biogeography of the cristata group is currently summarized as follows. Of the six Colocasiomyia species groups, the cristata group is most widely distributed in the Oriental and Papuan regions, covering nearly the entire range of the genus Colocasiomyia from Solomon Islands / Papua New Guinea (east) to Sri Lanka (west) and from Java (south) to Ryukyu Islands/ Taiwan /South China (north) ( Fig. 14 View FIGURE 14 ). The cristata and colocasiae subgroups coexist almost across this range. The three species complexes of the cristata subgroup are more or less different in their distribution ranges. The sabahana complex is endemic to Borneo, and the cristata complex is distributed in the Greater Sunda Islands and Philippines. The xenalocasiae complex is most widely distributed from tropical to subtropical areas of the whole Colocasiomyia range; C. xenalocasiae is confined to the subtropical area, where it shares the same host plant with C. alocasiae of the colocasiae subgroup ( Honda-Yafuso 1983; Yafuso, 1994; Toda unpublished data). Of the three species unassigned to any species complex, C. ecornuta and C. vieti are endemic to Sulawesi and Vietnam, respectively, and C. grandis cohabits in the same host-plant inflorescences with C. xenalocasiae and C. alocasiae in northern Vietnam and southern Yunnan ( Sultana et al. 2006).
Collection records of all species of the cristata group from their host plants at various localities in the Oriental and Papuan regions are compiled in Appendix 8. Modes of host-plant use vary from monopolization by a single species to sharing by 12 species. In ecological observations of this study, we focused on four species of the cristata subgroup, C. cristata and C. sulawesiana of the cristata complex, C. leucocasiae of the xenalocasiae complex and C. sabahana of the sabahana complex, each of which monopolizes its specific host plant.
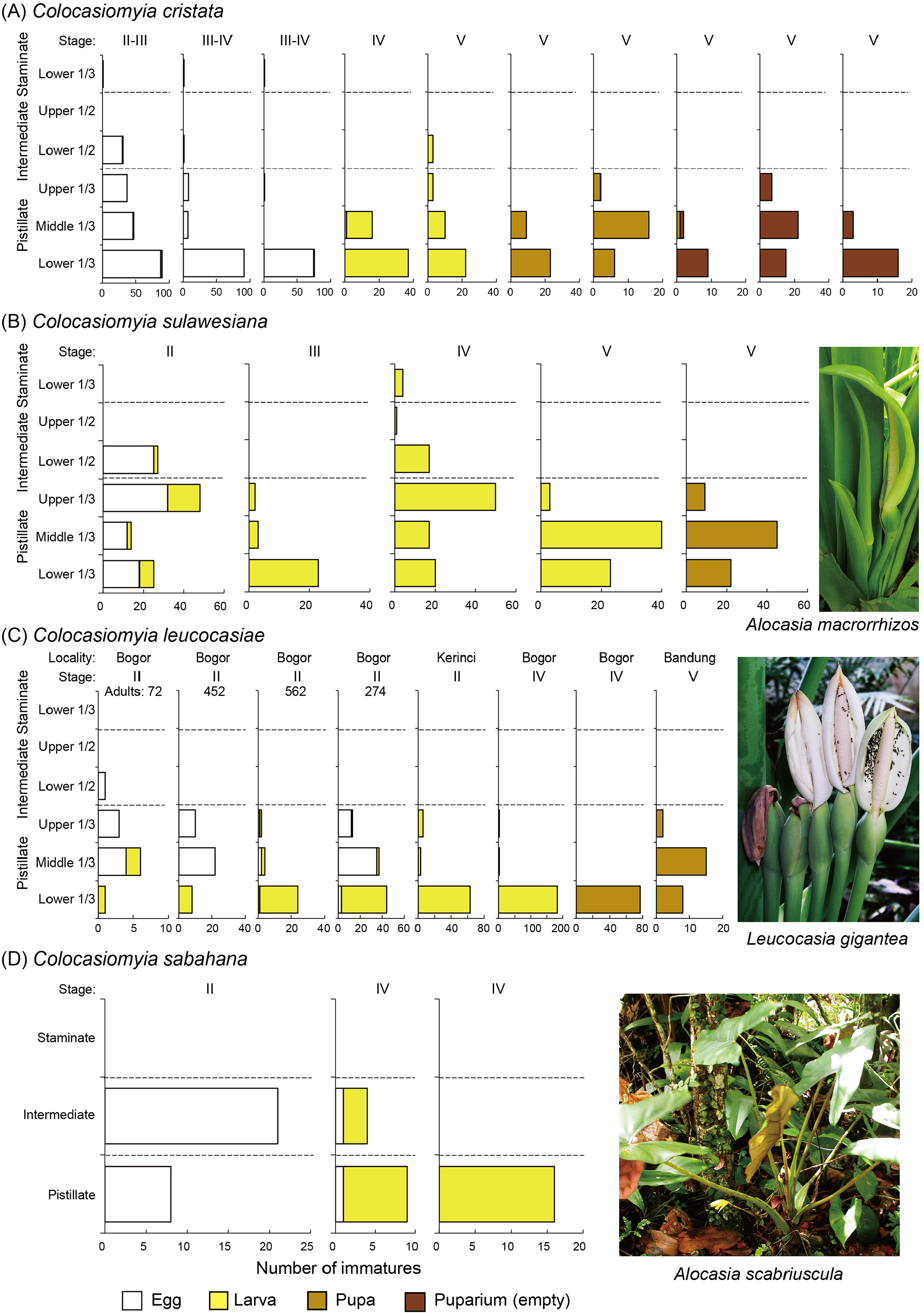
Colocasiomyia cristata was observed using inflorescences/infructescences of Alocasia alba in West Java. When the anthesis started with spathe opening and strong odor emission, adult flies came to the inflorescence at the female flowering phase (Stage II) and stayed in the spathal chamber around the pistillate and the lower intermediate regions of spadix, probably feeding, ovipositing and/or mating there ( Fig. 30A View FIGURE 30 ). Then, when the spathe began to constrict at the middle portion of intermediate region and pollen release started in the staminate region, the flies crawled up from the spathal chamber to the open staminate region of spadix ( Fig. 30B View FIGURE 30 ). Eggs were laid mostly on the pistillate region, especially its lower 1/3 section, sporadically on the lower 1/2 intermediate section, and very rarely on the lower 1/3 staminate section ( Figs 30C View FIGURE 30 , 31A View FIGURE 31 ). Larvae fed in these spadix portions of infructescences at Stages IV and V, and pupariated within the infructescences of Stage V ( Fig. 31A View FIGURE 31 ). Offspring adults emerged and left the infructescences through a hole at the top of spathal wall ( Fig. 30D View FIGURE 30 ) before the spathal dehiscence.
Colocasiomyia sulawesiana monopolized inflorescences/infructescences of Alocasia macrorrhizos in South Sulawesi. Eggs were laid on the pistillate region and the lower 1/2 intermediate section ( Fig. 31B View FIGURE 31 ). Young (1st and 2nd instar) larvae were found feeding there and rarely on the lower 1/3 staminate section of spadix in inflorescences/ infructescences at Stages II to IV ( Fig. 31B View FIGURE 31 ). Older (2nd and 3rd instar) larvae were found exclusively from the pistillate region, most abundantly from its middle 1/3 section, in infructescences of Stage V and pupariated there ( Fig. 31B View FIGURE 31 ).
Adult flies of C. leucocasiae sp. nov. were collected, often together with those of C. gigantea , from inflorescences of Leucocasia gigantea at the Bogor Botanical Garden in West Java. However, Fartyal et al. (2013) revealed that newly eclosed, young C. gigantea flies visit inflorescences of this host plant for feeding, and that the mature flies use inflorescences of Epipremnum pinnatum (L.) Engler (Monsteroideae, Araceae ) for breeding. Thus, C. leucocasiae sp. nov. monopolizes inflorescences/infructescences of Leucocasia gigantea for breeding at the Bogor Botanical Garden. Eggs were laid mainly on the pistillate region, most abundantly on its middle 1/3 section, but rarely on the lower 1/2 intermediate section ( Fig. 31C View FIGURE 31 ). Larvae were confined to the pistillate region, mostly its lower 1/3 section, and pupariated on the lower pistillate region of infructescences at Stage IV ( Fig. 31C View FIGURE 31 ).
Colocasiomyia sabahana sp. nov. monopolized inflorescences/infructescences of Alocasia scabriuscula in the headquarters area of Kinabalu Park, Sabah. Eggs were laid on the pistillate and intermediate regions, more on the latter region, and young (1st and 2nd instar) larvae were found feeding there, more in the former region of inflorescences at Stage IV ( Fig. 31D View FIGURE 31 ). At Stage V, the dried appendix falls off but the decayed staminate region remains within the spathal capsule of infructescence. Third-instar larvae were observed feeding on the pistillate and staminate regions of some infructescences at Stage V, and empty puparia were found from other Stage-V infructescences, though the numbers of individuals of these pre-imaginal stages were not counted.
Thus, all these four species of the cristata subgroup, each of which monopolizes its own host plant for breeding, are pistilicolous, i.e. ovipositing, feeding (larvae) and pupariating mostly on the pistillate (lower, female-flowers) region of spadix, as well as three species ( C. pistilicola , C. diconica and C. xenalocasiae ) of the xenalocasiae complex, each of which shares its host plant with another stamenicolous species ( C. stameniicola , C. colocasiae and C. alocasiae , respectively) of the colocasiae subgroup ( Carson & Okada 1980; Toda & Okada 1983; Honda-Yafuso 1983; Yafuso, 1994). In addition, two cristata- subgroup species, C. kotana sp. nov. and C. kinabaluana sp. nov., which share Alocasia macrorrhizos as their common host plant, are both pistilicolous as well, with slight breedingniche differentiation ( Takano et al. 2012).
Based on the host-use records compiled in Appendix 8, the cristata group is regarded as specializing on the following four genera of the subfamily Aroideae : Alocasia (11 spp.), Colocasia (7 spp.), Leucocasia (1 sp.) and Steudnera (1 sp.). To estimate evolutionary changes and/or acquirement of host plants at the generic level in the cristata group, the four host-plant genera were mapped on the phylogenetic tree having resulted from the morphological grafting cladistic analysis. ACCTRAN and DELTRAN resulted in the same inference of ancestral states ( Fig. 32 View FIGURE 32 ). All the species of the gigantea group (outgroup) use exclusively monsteroid plants for breeding ( Fartyal et al. 2013; Li et al. 2014; Jiao et al. 2020). The most recent common ancestor (MRCA) of the cristata group was estimated to have acquired Alocasia as its host plant. Nine extant species of the cristata subgroup use solely Alocasia for breeding with pistilicolous habits so far as known (five of the nine species). Therefore, the MRCA of the cristata group would have been pistilicolous as well. Of the nine Alocasia specialists, eight species are monoxenous, each depending on a single host-plant species; Alocasia macrorrhizos is the most important host plant, being used by four of them (Appendix 8). But, C. sarawakana sp. nov. is polyxenous, having been recorded from four Alocasia species (Appendix 8).
After the acquirement of Alocasia as host plant by the MRCA of the cristata group, host change from Alocasia to Colocasia was estimated to have occurred on two branches, i.e. in the MRCAs of the colocasiae subgroup and of the xenalocasiae complex; all the descendant, extant species of these ancestors are known to visit Colocasia inflorescences. Three pairs each consisting of one stamenicolous colocasiae- subgroup species and one pistilicolous xenalocasiae- complex species cohabit in the same host inflorescence: C. stamenicola and C. pistilicola ( Carson & Okada 1980) , C. colocasiae and C. diconica ( Toda & Okada 1983) , and C. alocasiae and C. xenalocasiae (Honda- Yafuso 1983; Yafuso, 1994). Okada (1980) hypothesized that this cohabitation was established by a pair of ancestors of the colocasiae subgroup and of the xenalocasiae complex, and that such a pair dispersed and differentiated into the three extant pairs in different geographic areas. The cohabitation would have been achieved by micro-allopatric breeding-site differentiation along the spadix of host plant between two species of the ancestral pair: the ancestor of the colocasiae subgroup would have changed its breeding habits from pistilicolous ones to stamenicolous ones, while the ancestor of the xenalocasiae complex would have retained pistilicolous habits. This change of breeding habits in the former may have been forced by the host-plant change or through interspecific competition with the cohabitant species. In this respect, C. kotana sp. nov. and C. kinabaluana sp. nov. of the cristata complex are interesting: they are both pistilicolous and cohabit in the same inflorescence/infructescence of the host plant Alocasia macrorrhizos ( Takano et al. 2012) . However, C. kinabaluana sp. nov. tends to be more stamenicolous than C. kotana sp. nov., having the following traits: the longer oviscapt ( Figs 21F View FIGURE 21 , 22F View FIGURE 22 ), a larger aggregation of adult flies in the upper part of spathal chamber and more eggs deposited on the intermediate region. This situation may or may not correspond to the initial phase of cohabitation of the colocasiae -subgroup and xenalocasiae -complex ancestors. Another possible case of host-plant change is of C. leucocasiae sp. nov. Although only one male specimen of this species has been collected, together with C. diconica , C. sumatrana sp. nov., C. colocasiae and C. iskandari , from an inflorescence of Colocasia esculenta in West Sumatra, this species is very abundantly collected from inflorescences of Leucocasia gigantea ( Fig. 31C View FIGURE 31 ) and monopolizes it for breeding. Thus, this species may have changed its host plant from Colocasia to Leucocasia .
Acquirement of additional host-plant genera, i.e. niche expansion, was estimated to have occurred in four species ( Fig. 32 View FIGURE 32 ). Of the three host-sharing pairs, the pair of C. alocasiae and C. xenalocasiae expanded their host plants to Alocasia and Leucocasia in their northernmost distribution area (subtropical zone), probably due to the scarceness of Colocasia inflorescences there. In addition, C. grandis sp. nov. joined this pair and formed a particular system of three-species cohabitation in a small area restricted to northern Vietnam and southern Yunnan. Within the colocasiae subgroup, which principally depends on Colocasia , C. steudnerae has acquired Steudnera colocasiifolia as another host plant and monopolistically uses its inflorescences/infructescences for breeding ( Takenaka et al. 2006). Interestingly, probably in connection with the monopolization of this new host plant, C. steudnerae exhibits a mixture of pistilicolous and stamenicolous breeding habits: ovipositing exclusively on the pistillate region (pistilicolous habit), larvae feeding on decayed tissues of the staminate region (stamenicolous) and pupariating out of the infructescence (stamenicolous). Furthermore, a morphological adaptation for the first pistilicolous habit is seen in the oviscapt: C. steudnerae has the distally broad oviscapt, as well as other pistilicolous species of the cristata subgroup, which seems to be adaptive for laying eggs between pistils placed more loosely. In contrast, other stamenicolous species of the colocasiae subgroup have the distally very narrow oviscapts to insert their eggs into narrow space between stamens packed compactly. With regard to the evolution of these mixed habits and morphology, Takenaka et al. (2006) proposed two hypotheses. The first is that C. steudnerae has secondarily evolved the pistilicolous oviposition habit and the related morphology of oviscapt under the condition lacking a pistilicolous cohabitant species on the new host plant Steudnera colocasiifolia . The second hypothesis is that the mixed situation in C. steudnerae represents an initial, transitional phase of evolution from ancestral, pistilicolous habits to stamenicolous ones in the colocasiae subgroup. The results of phylogenetic analyses in the present study lend support to the first hypothesis. However, more evidence is needed to fully depict the evolution of breeding habits in the colocasiae subgroup: a total of 14 undescribed species of this subgroup, most of which are sympatric with C. steudnerae in northern Vietnam and southern Yunnan ( Fig. 14 View FIGURE 14 ), are awaiting taxonomical, phylogenetical and ecological studies on them. Moreover, cohabiting mechanisms in some systems where more than two Colocasiomyia species are incorporated have remained unexplored at all.






| MZB |
Museum Zoologicum Bogoriense |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Colocasiomyia leucocasiae Toda
| Takano, Kohei Takenaka, Gao, Jian-Jun, Hu, Yao-Guang, Li, Nan-Nan, Yafuso, Masako, Suwito, Awit, Repin, Rimi, Pungga, Runi Anak Sylvester, Meleng, Paulus Ak, Kaliang, Clement Het, Chong, Lucy & Toda, Masanori J. 2021 |
Colocasiomyia sp.1 aff. diconica:
| Takano, T. K. & Suwito, A. & Gao, J. J. & Yin, J. T. 2011: 22 |
| Sultana, F. & Hu, Y. G. & Toda, M. J. & Takenaka, K. & Yafuso, M. 2006: 694 |