
Juxtacribrilina ezoensis, Dick & Grischenko & Gordon & Ostrovsky, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5016.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:6E3BF843-16C1-4B91-AB72-D5C1D556384A |
|
persistent identifier |
https://treatment.plazi.org/id/312B8C48-E297-4377-B55B-11B8EA330130 |
|
taxon LSID |
lsid:zoobank.org:act:312B8C48-E297-4377-B55B-11B8EA330130 |
|
treatment provided by |
Plazi |
|
scientific name |
Juxtacribrilina ezoensis |
| status |
sp. nov. |
Juxtacribrilina ezoensis n. sp.
( Figs 4–8 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 ; Table 2)
urn:lsid:zoobank.org:act:312B8C48-E297-4377-B55B-11B8EA330130
Cribrilina annulata: Grischenko 1997, p. 171 View in CoL ; Grischenko et al. 2007, p. 1093, fig. 17; Yang et al. 2018, figs 11, 12.
Diagnosis. Non-ovicellate zooids with 12–18 costae (colony averages 12.8–15.2), 3 or 4 oral spines (predominantly 3); intercostal lacunae medium-sized, 5–9 in transverse series (colony averages 6.9–7.5). Most zooids with 1 or 2 frontal pore chambers on proximal gymnocyst. Ovicellate zooids present in basal layer in subtidal specimens, rare in basal layer in intertidal specimens; roughly same size (including ooecium) as non-ovicellate zooids. Reduced and dwarf ovicellate zooids occur in basal layer near or at colony margin. Frontally budded dwarf ovicellate zooids typically present, often tightly packed in colony center, sometimes few or lacking; frontal shield usually with 3 or 4 costae. Ooecium reduced, cap-like, kenozooidal, bearing several irregular pseudopores in ovicellate zooids in basal layer, typically two transversely arranged pseudopores in dwarf zooids. Ancestrula cribriform, with 4 oral spines.
Etymology. The specific name derives from Ezo , the old Japanese name for Hokkaido Island.
Material examined. Holotype: YPM-IZ-106549, Akkeshi Narrows , Akkeshi Bay, Hokkaido, Japan (43.048073°N, 144.85021°E), collected by M.H. Dick, July 2014, dredge, depth 6–8 m, encrusting dead bivalve shell; on SEM stub GoogleMaps . Paratypes: YPM-IZ-106550, YPM-IZ-106551, YPM-IZ-106552, same collection information as for holotype, on SEM stubs; YPM-IZ-106553 (CaAkk-1), Kakijima Island , Akkeshi-ko (=Akkeshi estuary), Akkeshi , Japan (43.04145°N, 144.86061°E), collected by A. V. Grischenko and M.H. Dick, 4 June 2004, rocky intertidal; YPM-IZ-106554, YPM-IZ-106555, NHM 2006.2.27.31, Aininkappu, Akkeshi Bay , Japan (42.99267°N, 144.85500°E), collected by A. V. Grischenko and M.H. Dick, 5 June 2004, rocky intertidal, on SEM stubs; NHM 2006.2.27.50, NHM 2006.2.27.51, Daikokujima I., Akkeshi Bay, Japan (42.95433°N, 144.87567°E), collected by A. V. Grischenko and M.H. Dick, 7 June 2004, rocky intertidal, on SEM stubs; NHM 2006.2.27.52, Barasan Cape, Akkeshi Bay, Japan (43.02417°N, 144.83683°E), collected by A. V. Grischenko and M.H. Dick, 3 July 2004, rocky intertidal, on SEM stub GoogleMaps . Other material: YPM-IZ-106556, East Tongass Narrows, Ketchikan, Alaska (55.31657°N, 131.51990°W), collected by M.H. Dick, 11–12 September 2003, rocky intertidal; ZIRAS 01 /50734, Bering Island ( Commander Islands ), Station 10, Cape Gaupta (Kitovyy) (55.24583°N, 165.92056°E), one colony encrusting barnacle fragment, collected by A GoogleMaps . V. Grischenko, 1 August 1991, lower horizon of rocky-boulder intertidal, Laminaria dentigera zone, rock face .
Measurements. See Table 2.
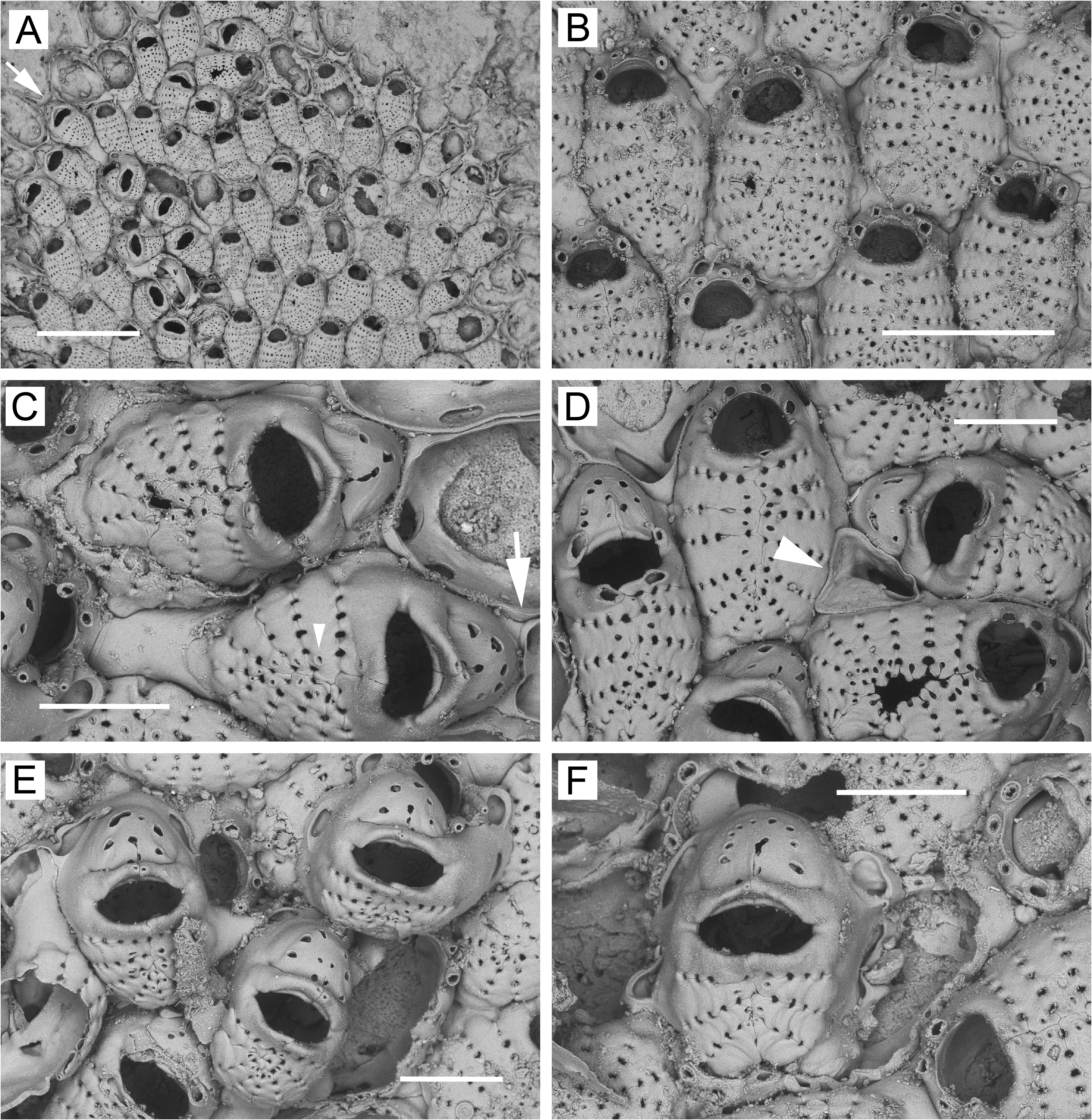
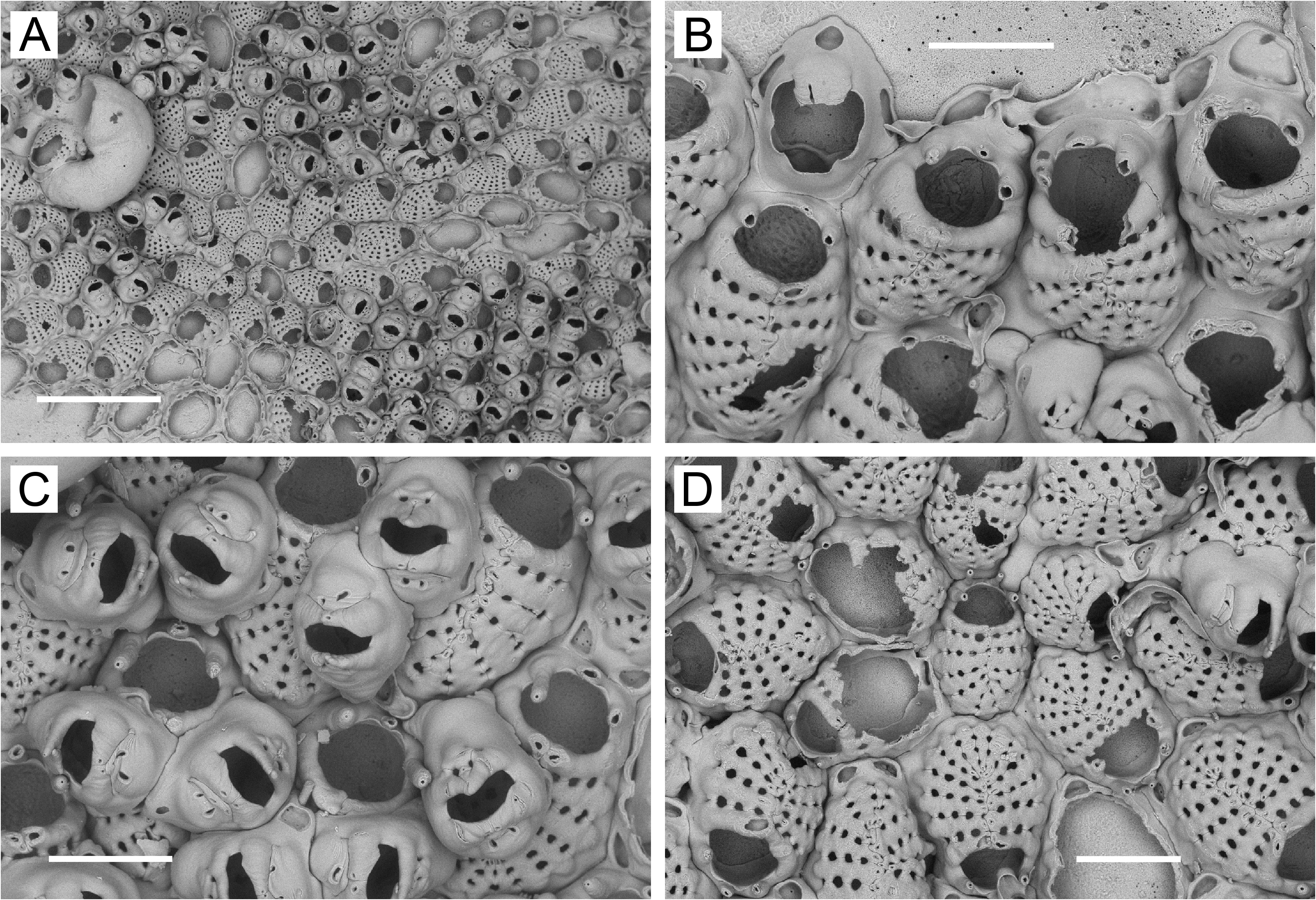
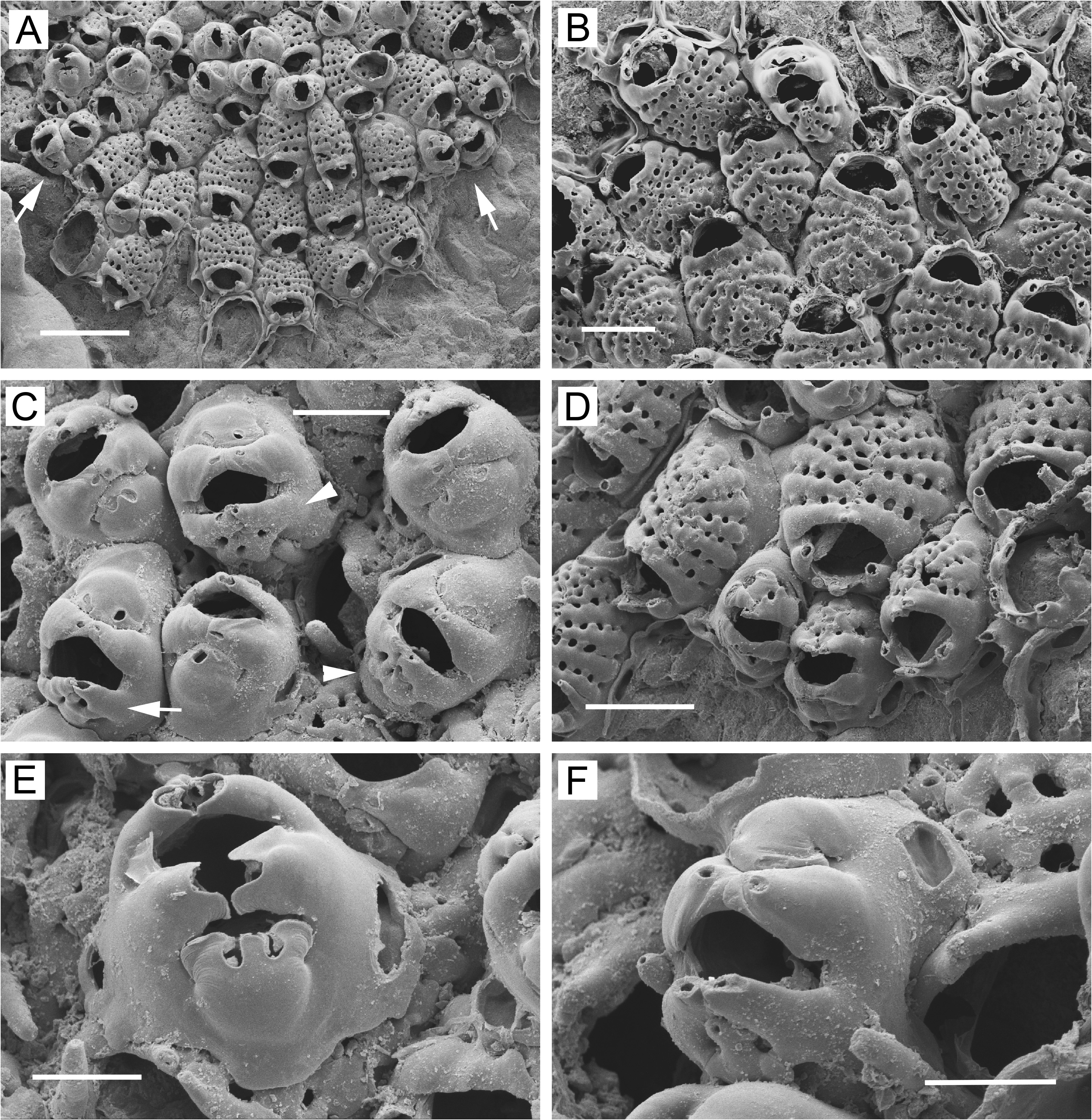
Description. Colonies encrusting, multiserial, unilaminar except for frontally positioned dwarf zooids ( Figs 4B, C View FIGURE 4 , 6A, B View FIGURE 6 , 7A View FIGURE 7 , 8A View FIGURE 8 ). Non-ovicellate zooids ( Figs 4A, B View FIGURE 4 , 5B View FIGURE 5 , 6B View FIGURE 6 , 7B View FIGURE 7 , 8B View FIGURE 8 ) oval or sometimes spindle-shaped, widest in middle, closely packed; gymnocyst negligible, sometimes wider in proximal corners or extending proximally as long caudal zone. Most ovicellate and non-ovicellate zooids in basal layer bearing 1 or 2 (occasionally 3) oval, triangular, or quadrangular frontal pore chambers on proximal gymnocyst ( Figs 4A, B View FIGURE 4 , 7B View FIGURE 7 , 8B View FIGURE 8 ); frontal pore chambers multiporous ( Figs 4E View FIGURE 4 , 5E View FIGURE 5 ). Frontal shield highly convex, sometimes slightly keeled; 12–18 costae (average values, 12.8–15.2) in non-ovicellate zooids; oval or somewhat flattened in cross section; suboral pair not appreciably wider than rest but sometimes somewhat swollen, often with small, fused tips extending fronto-distally ( Figs 4B, E View FIGURE 4 , 5B View FIGURE 5 , 8B View FIGURE 8 ); each costa with minute lumen pseudopore at or near tip. Intercostal lacunae moderate in size; circular, oval, subcircular, or heart-shaped (end closest to midline tapering); 5–9 in transverse series (average per colony, 6.9–7.5), most zooids with 6–8. Secondary orifice of non-ovicellate zooids roughly semicircular, wider than long, with corners widely rounded. Oral spines most commonly 3 (sometimes 4; rarely, 2 or 5), lateral pair often longer and heavier than spine(s) in between; single median spine usually flattened, widest basally, tapering distally, paired median spines usually cylindrical, occasionally fused.
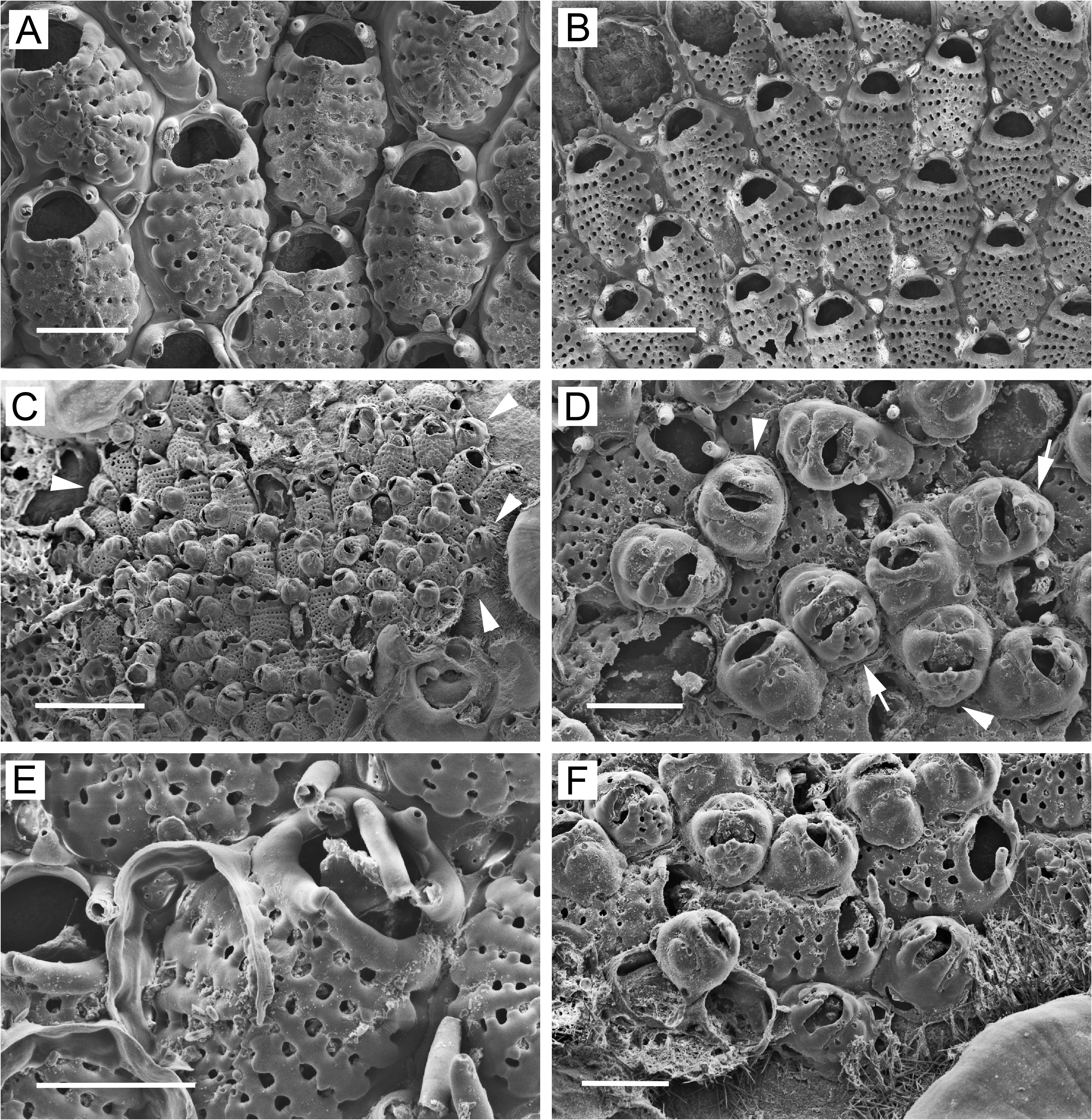
Ovicellate zooids lacking in basal layer (except at margin) in intertidal colonies but usually present in basal layer in subtidal colonies, either scattered individually ( Fig. 6B View FIGURE 6 ) or in circumferential bands ( Figs 5A–C View FIGURE 5 , 6A View FIGURE 6 ). Ovicellate zooids (including ooecium) in colony interior as large as non-ovicellate zooids ( Figs 5A View FIGURE 5 , 6B View FIGURE 6 ), with 11–17 costae; suboral pair often markedly wider than rest, with part of fused ends projecting fronto-distally to form mucro proximal to and overhanging orifice. Mucro sharp, with double or single tip, or wide, blunt, flattened. Reduced zooids (with as few as ~8 costae) and dwarf zooids sometimes occurring in basal layer at or near colony margin ( Figs 4F View FIGURE 4 , 6A View FIGURE 6 , 7B View FIGURE 7 , 8A, B, D View FIGURE 8 ). Narrow, concentric zone of somewhat to markedly reduced ovicellate zooids seen toward margin of one colony ( Fig. 6A View FIGURE 6 ). Ovicellate zooids in basal layer with distal pore chamber, sometimes nonbudding and evident as rounded opening below ooecium ( Fig. 5B View FIGURE 5 , arrow), but usually budding distal zooid ( Fig. 5B View FIGURE 5 , arrowheads); basal ovicellate zooids thus commonly occurring in columnar series with other zooids ( Figs 5A View FIGURE 5 , 6A View FIGURE 6 ).
Ooecium in basal zooids ( Figs 5B–D View FIGURE 5 , 6A View FIGURE 6 ) kenozooidal, arising from roof of distal pore chamber of maternal zooid ( Fig. 6D View FIGURE 6 ), connecting with pore chamber via two or more pores ( Fig. 6D, F View FIGURE 6 ); reduced, cap-shaped, abutting arching, modified pair of wide, flattened, non-articulated, latero-oral spines fusing at midline; modified spines tapering and twisted toward midline, forming distal convexity of secondary orifice; ends often flattened and raised to become blunt median projection. Secondary orifice shaped like smiling mouth flanked by lips (widened suboral costae and modified latero-oral spines). Ooecium of zooids in basal layer with 2–6 small to moderately large, scattered, circular to irregular pseudopores; proximal margin of cap not visible inside secondary orifice.
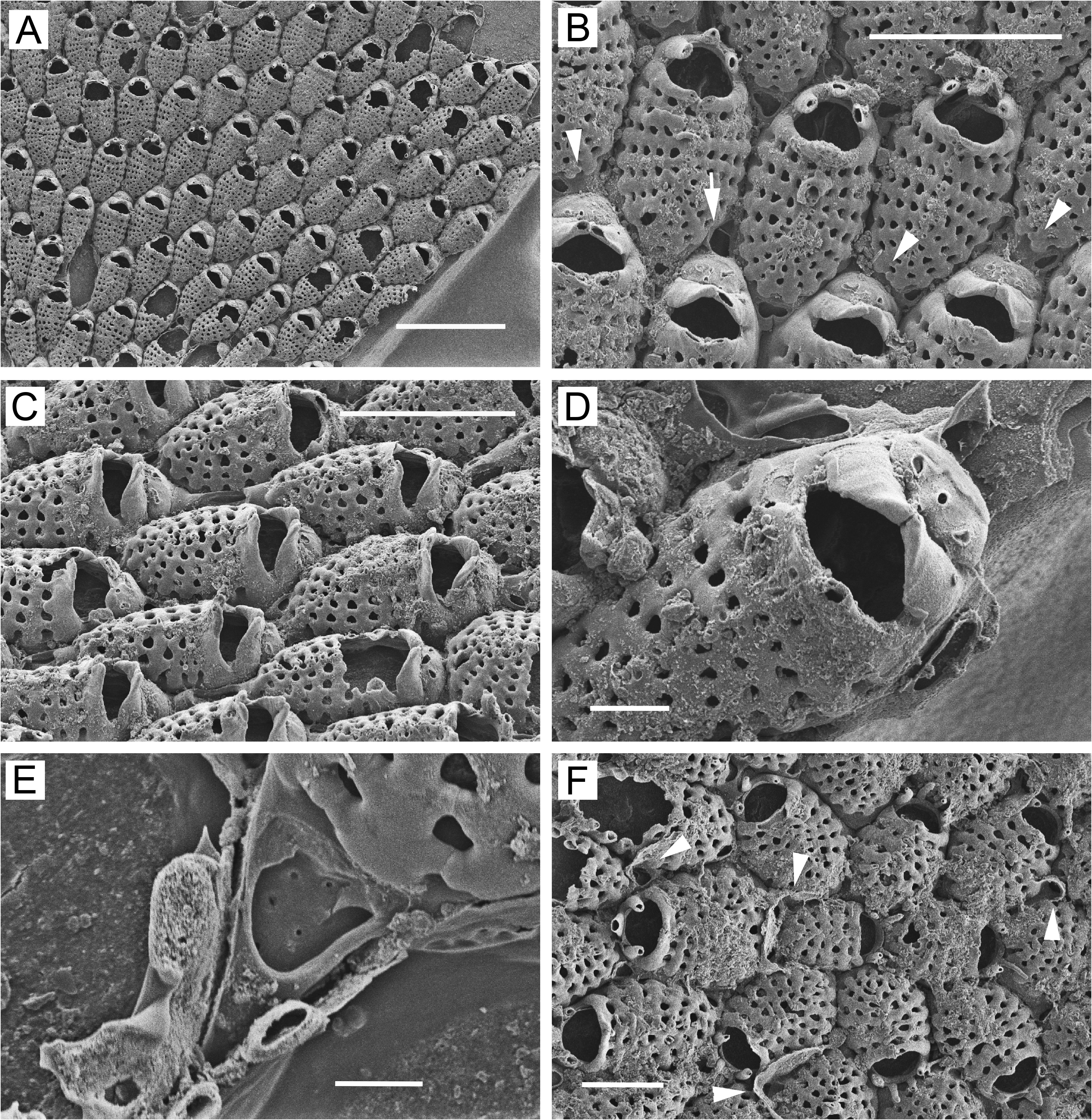
Dwarf zooids often abundant in center of colony ( Figs 4C View FIGURE 4 , 6B View FIGURE 6 , 7A View FIGURE 7 , 8A View FIGURE 8 ), budded from frontal pore chambers of basal zooids ( Fig. 4E View FIGURE 4 ); costal shield ( Figs 4D View FIGURE 4 , 6C View FIGURE 6 , 8C, F View FIGURE 8 ) reduced to 3 or 4 costae (occasionally 2 or 5) including suboral pair; suboral pair often curved in distal direction, forming sharp mucro or evenly tapering triangular process. Ooecium appears to be kenozooidal, budded distally from maternal zooid. Ooecial complex ( Figs 4D View FIGURE 4 , 6C View FIGURE 6 , 7C View FIGURE 7 , 8C, E, F View FIGURE 8 ) in dwarf zooids similar to that in ovicellate zooids in basal layer; modified latero-oral spines not twisted as much and usually not forming median process; cap-like ooecium similar but narrower, usually with 2 circular or oval, transversely arranged pseudopores ( Figs 4D View FIGURE 4 , 6C View FIGURE 6 , 7C View FIGURE 7 , 8C View FIGURE 8 ), these sometimes partly merged. Margin of ooecial cap meeting modified spines; sometimes slightly thickened and bearing 1 or 2 tiny additional pseudopores. Dwarf zooids sometimes with 1–2 basal pore chambers evident in distal (in ooecial kenozooid), disto-lateral, lateral, or proximo-lateral positions ( Figs 6C View FIGURE 6 , 8C, E, F View FIGURE 8 ). Marginal, basally budded dwarf zooids ( Figs 4F View FIGURE 4 , 8D View FIGURE 8 ) more common in intertidal than in subtidal colonies.
Ancestrula ( Fig. 5F View FIGURE 5 ) cribriform, similar to but smaller than subsequent zooids, with 4 articulated oral spines.
Remarks. Grischenko et al. (2007) described this species in detail (as Cribrilina annulata ) from the intertidal zone at Akkeshi Bay, Japan. Juxtacribrilina ezoensis n. sp. differs from J. annulata in several clear-cut ways. In J. ezoensis n. sp., most zooids in the basal layer have one or two frontal pore chambers on the proximal gymnocyst, whereas in J. annulata , frontal pore chambers occur only rarely and are usually entirely absent. Consequently, J. ezoensis n. sp. can readily bud frontal dwarfs at any time during colony astogeny, and in any part of the colony. In contrast, as far as is known, J. annulata produces frontal dwarfs only from basal pore chambers, which once a colony has formed are largely unavailable for budding in the center of the colony. Frontal dwarfs are therefore much more common in J. ezoensis n. sp. and often occur abundantly in the central region of the colony, whereas in J. annulata they are uncommon and usually occur at or near the colony margin.
The two species also differ markedly in the size of the ooecium and the form and structure of the ooecial complex. In J. annulata , the ooecium is hood-like and only moderately reduced from the full-sized ooecium seen in most cheilostomes ( Ostrovsky 1998, 2008, 2013, 2020; Martha et al. 2020); the modified latero-oral spines arching over the proximal part of the ooecium retain a spine-like appearance and do not obscure the proximal ooecial margin ( Figs 2D–F View FIGURE 2 , 3C, D View FIGURE 3 ). The ooecium is kenozooidal, budded from the distal area of the maternal zooid occupied by the septulum of the forming basal pore chamber; in both basal and dwarf zooids, the kenozooidal ooecium may itself give rise to one or two distal pore chambers, but only rarely ( Fig. 2C View FIGURE 2 ) has been observed to produce another autozooid distally. Ovicellate zooids in the basal layer thus do not usually bud a distal autozooid, and usually do not lie in columnar series with following zooids ( Fig. 2D View FIGURE 2 ).
In contrast, the ooecium in J. ezoensis n. sp. is reduced to a broad, low cap and is more highly integrated with the modified spines, which are flatter and broader, forming the transverse bar that comprises the proximal margin of the ooecial complex and whose concave proximal edge contributes to the shape of the secondary orifice ( Fig. 5B, D View FIGURE 5 ). The ooecium in basal zooids is kenozooidal, but arises from the roof of the distal pore chamber ( Fig. 6D View FIGURE 6 ), to which it is connected via one or more communication pores ( Fig. 6F View FIGURE 6 ), thus leaving the pore chamber free to produce another zooid distally ( Fig. 6E View FIGURE 6 ); basal ovicellate zooids are thus typically arranged in columnar series ( Figs 5A, B View FIGURE 5 , 6A, B View FIGURE 6 ). In the infrequent cases where a basal ovicellate zooid is not in columnar series, the distal pore chamber remains evident as an oval opening below the ooecium ( Fig. 5B View FIGURE 5 , arrow).
Most measurements for zooids in the basal layer overlap considerably between J. annulata ( Table 1) and J. ezoensis n. sp. ( Table 2), especially considering variation among the populations within each of these species. However, in White Sea J. annulata , dwarfs are larger for all measurements and have more costae (range 4–7, average 5.7) than dwarfs in J. ezoensis n. sp. (overall range 2–5; range of averages 3.2–3.7). Ooecia in J. annulata have more pseudopores, both in basal ovicellate zooids (Kamchatka, 5–10; White Sea, 5–7) and in frontal dwarfs (White Sea, 3–5), than J. ezoensis n. sp. (basal ooecia, 2–6; dwarf ooecia, usually 2).
At Akkeshi, subtidal colonies of J. ezoensis n. sp. have "full-sized" ovicellate zooids in the basal layer, whereas intertidal colonies lack them ( Grischenko et al. 2007). Upon examination of additional intertidal material from Akkeshi, we again saw no "full-sized" ovicellate zooids in the basal layer, but did find dwarfs to occur in the basal layer at the colony margin ( Figs 4C, F View FIGURE 4 )—as was also the case for intertidal specimens of J. ezoensis n. sp. from the Commander Islands ( Fig. 7B View FIGURE 7 ) and Ketchikan ( Fig. 8A, D View FIGURE 8 ). While the differences in the intra-colony complement of ovicellate zooids between intertidal and subtidal material might seem to indicate two distinct species, we believe that the two forms are conspecific. The measurements for non-ovicellate and dwarf zooids ( Table 2) are remarkably similar between intertidal and subtidal specimens from Akkeshi, as are the number of intercostal lacunae in nonovicellate zooids, the form of the ooecium, and the form of the costal shield in dwarf zooids.
Previous studies have found the distribution of various types of ovicellate zooids (frontal or basal; full-sized or dwarf) in J. annulata colonies ( Nekliudova et al. 2019), and of zooids having different types of frontal shield ( Ito et al. 2015) in J. mutabilis , to be related to life history. In the case of J. ezoensis n. sp. at Akkeshi, we speculate that the paucity of ovicellate zooids in the basal layer in intertidal colonies is related to the less stable, more stressful intertidal environment compared to the subtidal environment. Subtidal colonies of J. ezoensis n. sp. at Akkeshi typically reach larger sizes than intertidal colonies and almost certainly overwinter. It may be that intertidal colonies, on the other hand, settle each season from subtidally produced larvae and do not overwinter, foregoing the production of larger ovicellate zooids in the basal layer in favor of frontal and marginal dwarfs. Frontal dwarfs typically appear first toward the colony center and are added centrifugally as the colony grows, presumably as conditions are favorable, allowing flexibility in the timing of reproduction and the resources devoted to it. Marginal dwarfs likely appear late in the season, as colony growth begins to wind down.
Among the Pacific Juxtacribrilina species treated in the subsequent descriptions, J. ezoensis n. sp. is most similar to J. pushkini n. sp. described next; a comparison of the two is deferred to the Remarks section for the latter.
Occurrence. J. ezoensis n. sp. has a trans-Pacific distribution; it was common intertidally and subtidally at Akkeshi Bay, Japan, and occurred intertidally in the Commander Islands (one colony) and at Ketchikan, Alaska (one colony).


| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Juxtacribrilina ezoensis
| Dick, Matthew H., Grischenko, Andrei V., Gordon, Dennis P. & Ostrovsky, Andrew N. 2021 |
Cribrilina annulata: Grischenko 1997 , p. 171
| Grischenko, A. V. & Dick, M. H. & Mawatari, S. F. 2007: 1093 |
| Grischenko, A. V. 1997: 171 |