
Ingolfiella alba, Iannilli, Valentina, Berera, Raffaella & Cottarelli, Vezio, 2008
|
publication ID |
https://doi.org/10.5281/zenodo.180292 |
|
DOI |
https://doi.org/10.5281/zenodo.6236184 |
|
persistent identifier |
https://treatment.plazi.org/id/506F87A9-FFA3-FFDC-FF35-59720AB96EA0 |
|
treatment provided by |
Plazi |
|
scientific name |
Ingolfiella alba |
| status |
sp. nov. |
Ingolfiella alba sp. nov.
Material examined. Holotype: female 1.85 mm, dissected and mounted on 3 slides labeled:
Ingolfiella alba , Marinduque, White Beach, 92-08-26, (MVRCr 448). Deposited in Museo Civico di Storia Naturale di Verona. Allotype: male 1.76 mm, dissected and mounted on 3 slides labeled: Ingolfiella alba , Marinduque, White Beach, 04-02-26, (MVRCr 442). Paratypes: one male dissected and mounted on slides labelled: Ingolfiella alba , Marinduque, White Beach, 04-02-26; 4 females dissected and mounted on slides labelled: Ingolfiella alba , Marinduque, White Beach, 92-08-26, numbered from 1 to 4; 2 females dissected and 3 females undissected and mounted on slides labelled: Ingolfiella alba , Marinduque, White Beach, 04-02- 26, numbered from 5 to 9; 2 females dissected and mounted on slides labelled: Ingolfiella alba , Oriental Mindoro, Long Beach,, numbered 10 and 11, respectively.
All material collected by V. Cottarelli
Diagnosis. Ocular lobes developed. Maxilla 1, outer plate with seven spines. Oostegites observed in pereopods 3–5. Gnathopods 1–2 carposubchelate, dactyli with four spiniform processes. Pereopods 3–4 with cilindrical trifid unguis. Pereopods 5–7 with dactyli similar, distally ending with bifid unguis. Pleopods 1–3 present, subtrapezoidal (subtriangular) similar. Uropod 1 with outer ramus length about two thirds of inner ramus; uropod 2 peduncle with three oblique rows of spines apically complex and long setae.
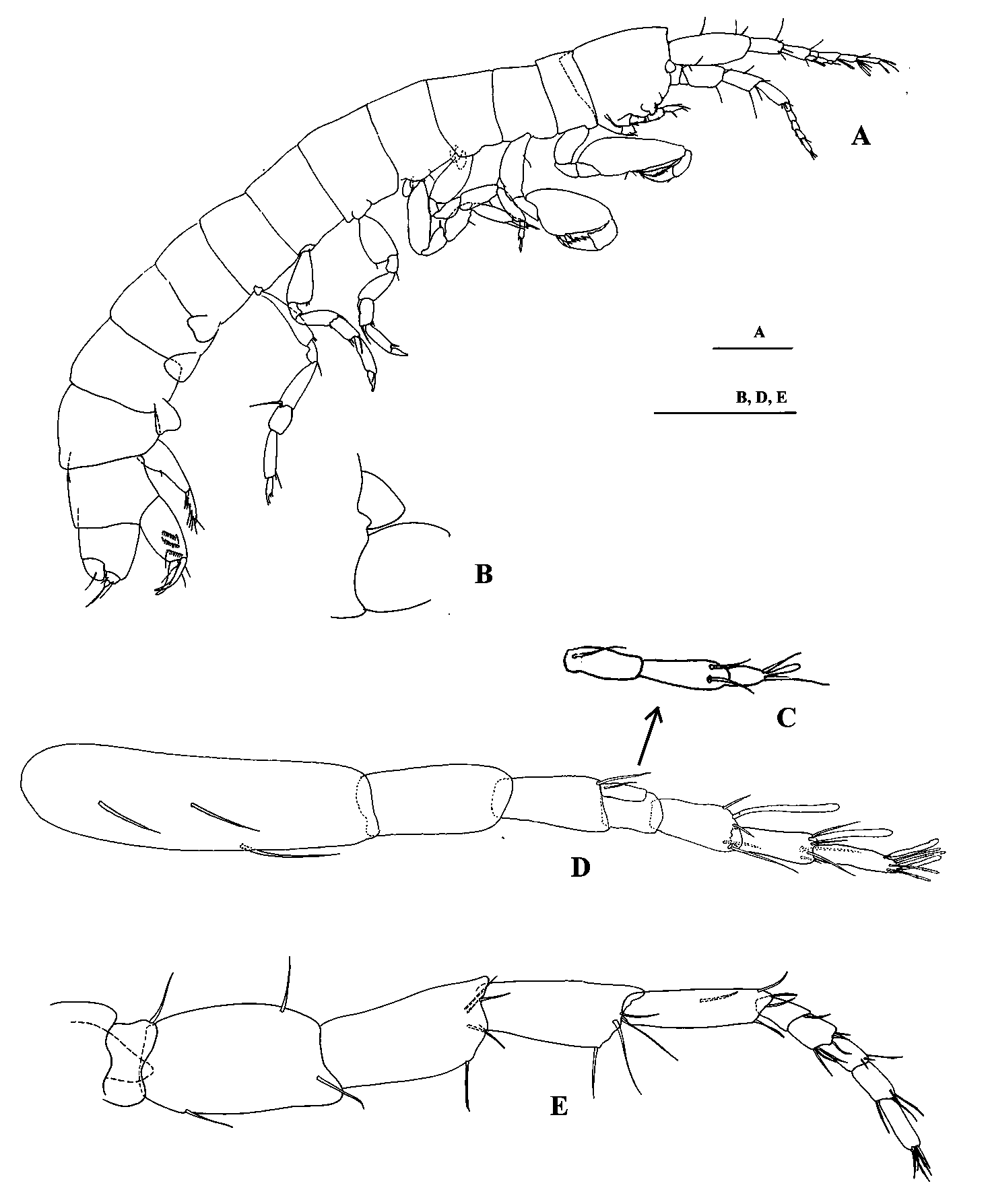
Description. Body elongate ( Fig. 1 View FIGURE 1 A), all segments laterally compressed. Head with weakly protruding rostrum; ocular lobe subtriangular ( Fig. 1 View FIGURE 1 A, B), reaching half length of the first article of antenna 2. Antenna 1 ( Fig. 1 View FIGURE 1 D), peduncular article 1 longer than articles 2+3; article ratio: 1: 0.4: 0.32; flagellum 4– articulate (first minute), shorter than peduncle, articles 2–4 with one or two aesthetascs; accessory flagellum ( Fig. 1 View FIGURE 1 C) 3-articulate, shorter than flagellar articles 1+2.
Antenna 2 ( Fig. 1 View FIGURE 1 E), peduncle longer than peduncle of antenna 1; flagellum 5-articulate, shorter than half length of peduncle, article 5 with one apical aesthetasc.
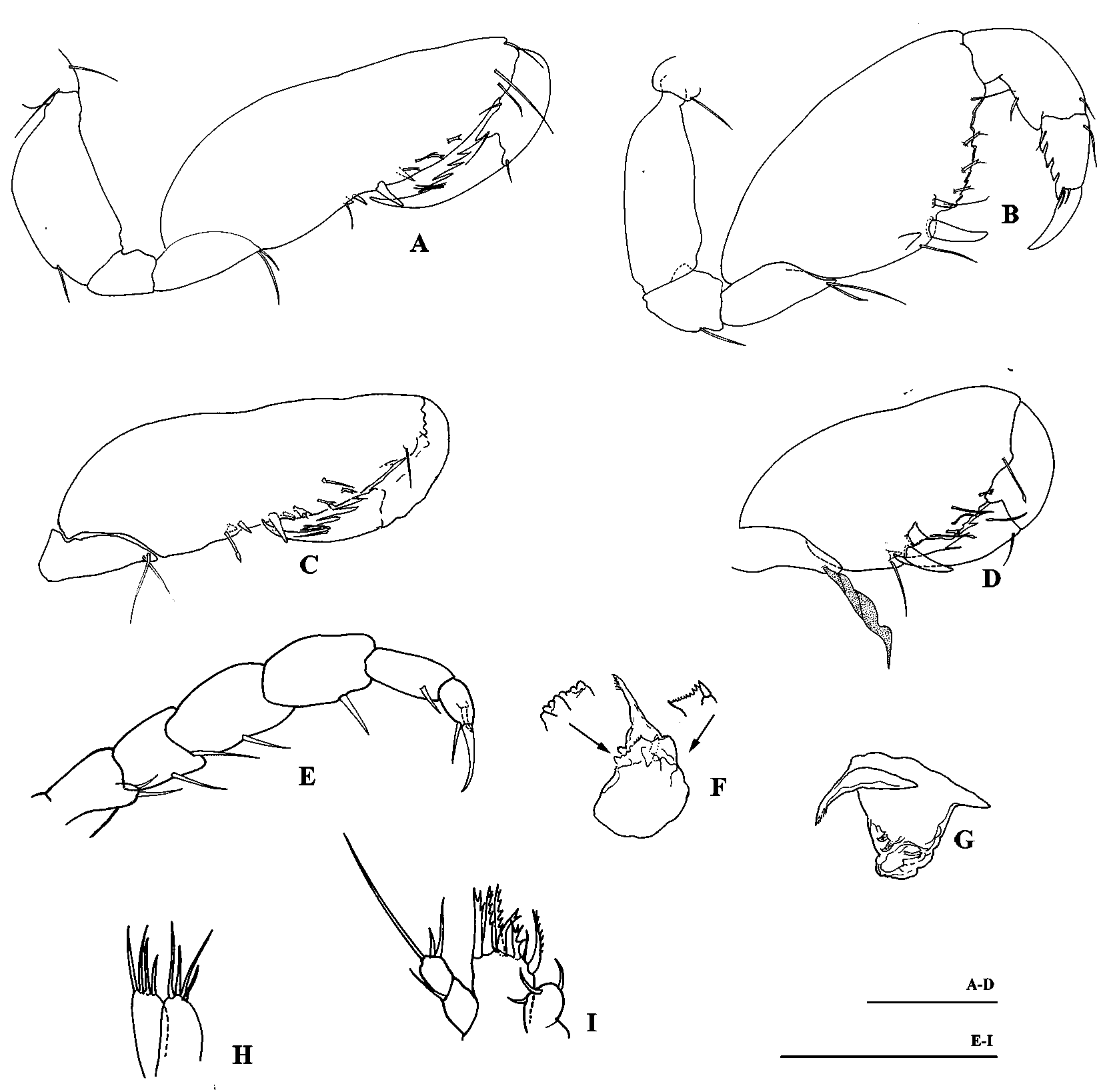
Mandible ( Fig. 2 View FIGURE 2 F, G) without palp, incisor with seven teeth, lacinia mobilis distally denticulate ( l. m. as broad as incisor) with several small teeth flanked by two large curved teeth, molar long and pointed, with minutely denticulate distal margin.
Maxilla 1 ( Fig. 2 View FIGURE 2 I), palp 2-articulate, just longer than (or subequal) outer plate, with three apical setae; outer plate with one 3-cuspidate, one 4-cuspidate and one 2-cuspidate spines on posterior row; two 6-cuspidate spines, strongly curved and one combed spine on anterior row.
Maxilla 2 ( Fig. 2 View FIGURE 2 H), inner and outer plate both with 4 naked apical setae.
Maxilliped ( Fig. 2 View FIGURE 2 E), basis endite bearing 2 apical setae, palp with 5 articles; palp articles 1–4 with one mesial seta each, palp article 2 with also one basal seta, article 5 (dactylus) with one strong spine and a long falcate unguis.
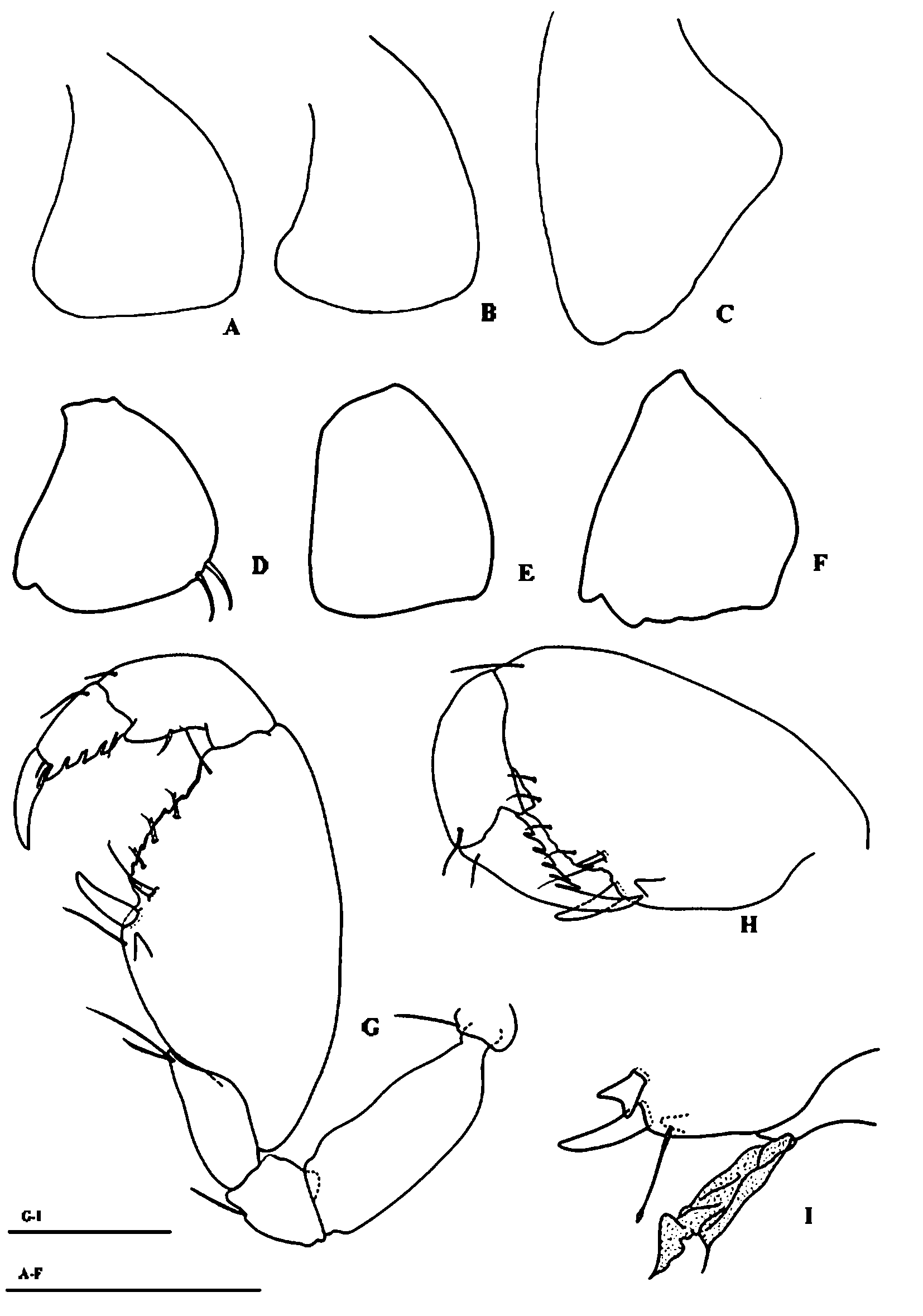
Gnathopod 1 ( Fig. 2 View FIGURE 2 A, C) carposubchelate, carpus relatively elongate and strong, inner face with 2 distal setae, anterior margin with 1 distal seta; propodus strong, with 2 setae on posterior and 1 on anterior margin; dactylus with 4 spiniform processes; palm strongly oblique with 4 setae apically bifid and one simple, 2 submedial spines (one of them strong) on inner face and one submedial tooth and one spine on outer face.
Gnathopod 2 ( Fig. 2 View FIGURE 2 B, D) carposubchelate, stronger than in gnathopod 1, c/p index (see Stock, 1977) =2.1; palm oblique, defined by a strong spine, with four or five setae bifid apically and one submedial spine, one seta (apically simple) and one tooth proximal to palmar corner spine, and margin with six or seven serrations; propodus strong, with one triangular blade and one distal seta posteriorly; dactylus with four or five ( Fig. 2 View FIGURE 2 B, D; Fig. 4 View FIGURE 4 G, H) subquadrate teeth on posterior margin and one simple proximal seta on anterior margin; Claw (unguis) elongate, overreaching the palmar spine. In male ( Fig. 2 View FIGURE 2 D; Fig. 4 View FIGURE 4 I), posterior margin, near to the palmar corner, with a heavy forked tooth, proximal to definitive palmar spine. Only in male specimens it is also present a foliaceus structure on the posterior margin of the merus.
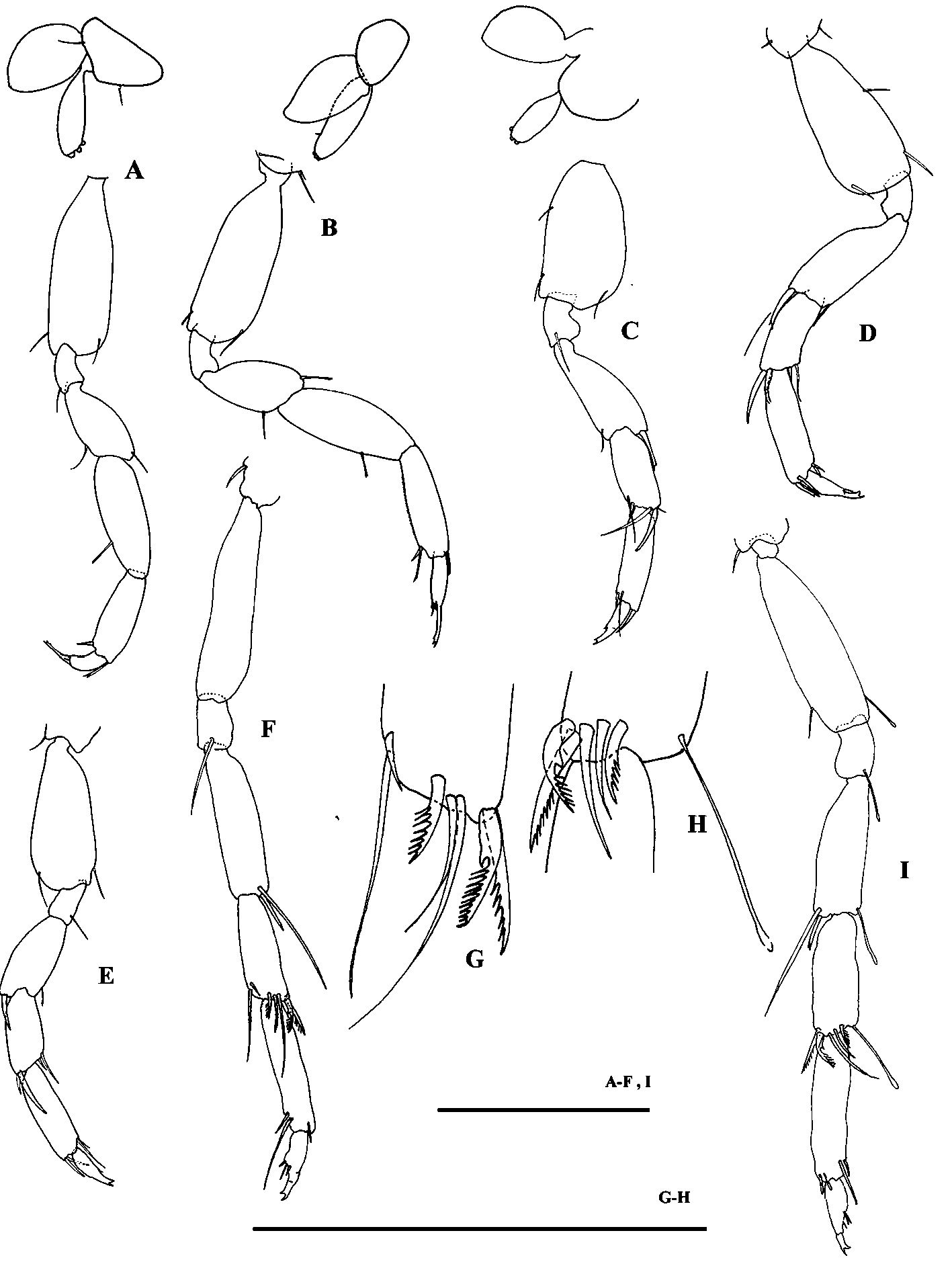
Pereopods 3–4 ( Fig. 3 View FIGURE 3 A, B) similar; pereopod 4, propodus and dactylus with many short setae on posterior margin, three distal spines on propodus, two of them apically bifid; dactylus length about half of propodus, distally bearing one or two setae apically complex, one spine, one seta and a slender and cylindrical trifid unguis.
Oostegites (on pereopods 3–5) small, suboval and elongate ( Fig. 3 View FIGURE 3 A, B, C), sometimes with one short seta; the main character is the presence of three bud-like (or button) processes.
Pereopod 5 ( Fig. 3 View FIGURE 3 C) shorter than pereopod 6, basis broad and short, heavy spines and setae on carpus and propodus; merus with one posterodistal bifid spine and one seta on anterodistal margin; dactylus similar to those of pereopods 6–7, feebly separated from unguis bifid, distally ending in two small spines.
Pereopod 6 ( Fig. 3 View FIGURE 3 D, E), basis longer and slightly narrower than P5; merus, carpus and propodus bearing many heavy spines (both simple and apically bifid); a very feeble demarcation line separates dactylus from unguis bifid.
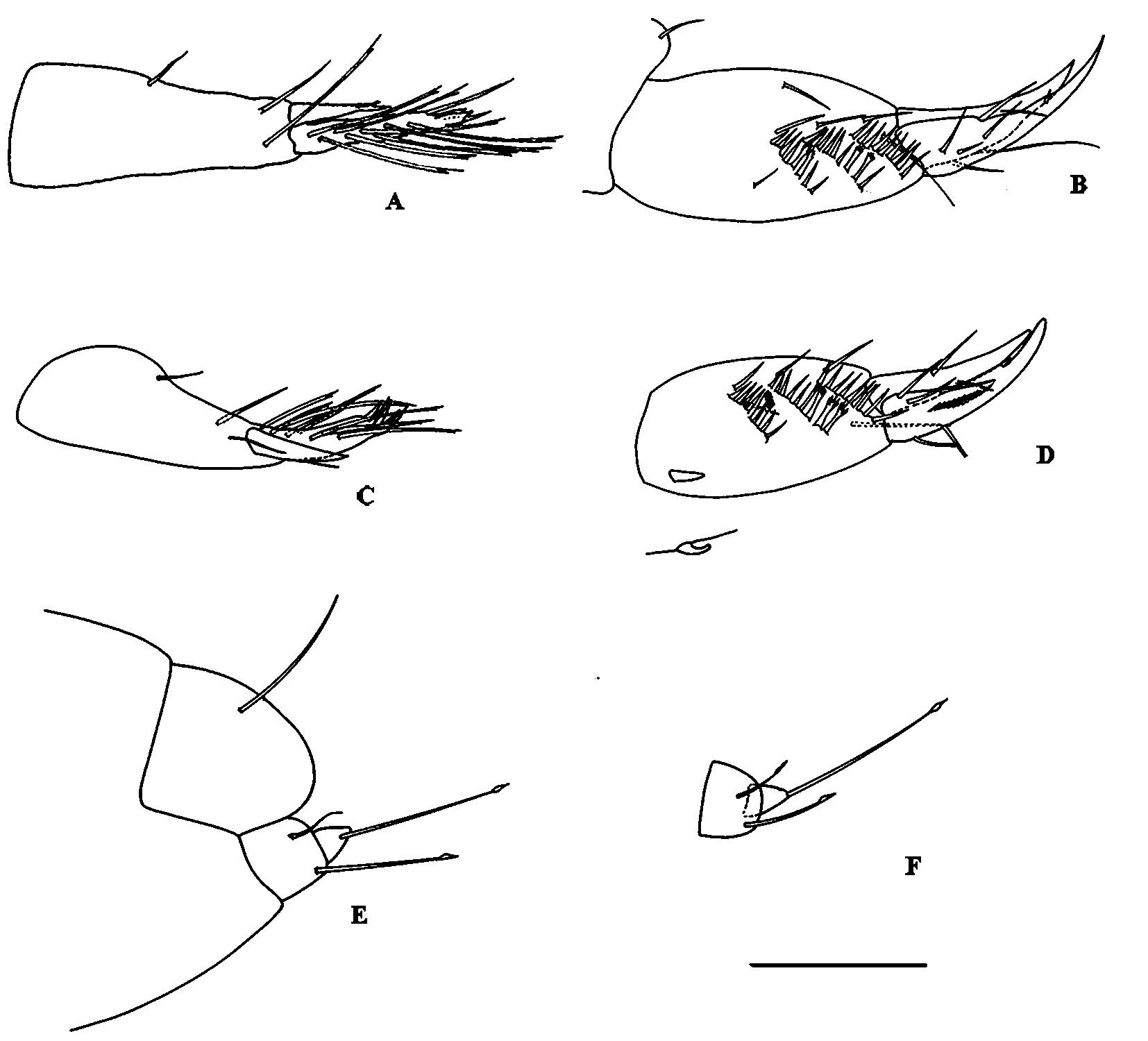
Pereopod 7 ( Fig. 3 View FIGURE 3 F, I) distinctly longer than pereopods 5–6; basis slender and elongate than in pereopods 5–6; merus with two spines (one of them very long) on posterodistal corner; carpus ( Fig. 3 View FIGURE 3 G, H) with many slender spines (shorter in male) and three strong modified (comb-like) spines; dactylus feebly separated from unguis bifid.
Pleopods 1–3 ( Fig. 4 View FIGURE 4 A, B, C) subtrapezoidal, similar, without setae; pleopod 3 slightly larger than pleopods 1–2. Pleopods in male ( Fig. 4 View FIGURE 4 D, E, F) with 1 or 3 lobes, pleopod 1 with 2 setae.
Uropod 1 ( Fig. 5 View FIGURE 5 A, C) biramous, peduncle longer than inner ramus, with two distal and one dorsomedial seta; outer ramus length about two thirds of inner ramus, distally pointed, with one seta; inner ramus, medial surface with eight or nine long setae and two distal spines on edge of denticulate apex with five or six spiniform processes.
Uropod 2 ( Fig. 5 View FIGURE 5 B, D), peduncle bearing three oblique rows, each of (proximal to distal) eight, nine, nine strong spines on inner face, some of them modified at tips (2+2+0 in female and 2+4+0 in male) and six setae; rami apically pointed (not in male); inner ramus, with five setae and one small distal spine; outer ramus shorter than inner one, with two setae near mid-length. It is also present a facial hooked spine ( Fig. 5 View FIGURE 5 D) in male specimens (that I. canariensis shows in female) in the proximal region of peduncle.
Uropod 3 ( Fig. 5 View FIGURE 5 E, F) very short, uniramous; peduncle with two long distal setae; ramus shorter than peduncle, broad, with one long distal seta.
Telson ( Fig. 5 View FIGURE 5 E) globose, with one pair of long dorsal setae.
Derivatio nominis. The specific name is derived from the Latin adjective “ alba ”, meaning “white”. It refers to White Beach, the location on Marinduque island where the types were collected, and to the white coral beaches where the new species was collected. These habitats are typical of coastal tropical areas, and are endangered by human impact (such as it happens in the collecting site in Mindoro). The epitheton is an adjective in feminine singular.
Affinities and remarks. The new species shows characters leading us to refer it to the Tethydiella group, the marine subgenus suggested by Ruffo & Vigna Taglianti (1989), with well developed ocular lobes, dactyli of pereopods 3–4 with claw elongated and apically trifid, unlike pereopods 5–7, with stout claw apically bifid. The authors attribute to this subgenus six marine species distributed from the Indian Ocean ( I. kapuri Coineau & Rao, 1973 ; I xarifae Ruffo, 1966 ); to the central Atlantic Ocean: Bermuda ( I. longipes Stock, Sket & Illfe, 1987 ); South Carolina and Florida ( I. fuscina Dojiri & Sieg, 1987 ); Caribbean region ( I. grandispina Stock, 1979 ; I. quadridentata Stock, 1979 ). It is also possible to recognise affinities with this subgenus in other species described after 1989: I. canariensis Vonk & Sanchez, 1991 , from Canary Islands; I. sandroruffoi Andres, 2004 from deep sea in the North Atlantic Ocean; the recently described species I. rocaensis Senna & Serejo, 2005 from a submerged atoll in Brazil and another Ingolfiella now being described from Arabian Sea (Abd el Kuri Island, Socotra Archipelago, Oman) by Iannilli & Ruffo.
Among the species of this group, Ingolfiella alba sp. nov. has the largest number of characters in common with I. xarifae described from Maldive Islands. We could analyse the type material of I xarifae preserved in Verona Natural History Museum. I. alba sp. nov. differs from I. xarifae mainly for the shape of carpus palm and posterior margin of dactylus of the gnathopod 2 with 5 serrations. In male a heavy forked spine near the palmar margin of the gnathopod 2 is present, as well as a foliaceous structure hanging from the posterior margin of the merus. This structure is also observed in I. canariensis and I. sandroruffoi . The c/p index is 2.1 in I. alba and 1.9 in I. xarifae . The new species differs also in pereopod 3, which shows the carpus longer than propodus and not the contrary as happen in I. xarifae . We can find also in I. alba sp. nov., several modified spines not described in I. xarifae and many other species, because it is not included in traditional characters and described only in recent descriptions.
The main differences among the tethydean species can be found in the shape and distance of spines and teeth on carpus palmar margin and serrations or teeth of gnathopod 2.
Ingolfiella alba sp. nov. presents a character never observed in other species of this genus: the oostegites present three small button-like processes. However, we don’t understand their significance. A character never observed in other species of this genus: something similar was described in Metaingolfiella mirabilis Ruffo, 1969 , although the processes were smaller and numerous. The re-examination of I. xarifae typus allowed us to observe also on the oostegites of this species the button-like processes described for I. alba . The same processes can be observed in the Ingolfiella now being described from Arabian Sea (Iannilli com. pers.). These structures are probably present in other Ingolfiella species but have yet to be observed and described.
Vonk & Schram (2003) recently proposed an important phylogenetic and biogeographic analysis of Ingolfiellidae , based on a 43 characters matrix. The genus Ingolfiella , was considered by those authors as “a single undivided taxon”, nevertheless the wide distribution area and the large variety of habitats occupied by this genus were not considered. Vonk & Schram (2003) did not accept the splitting of Ingolfiella in genus and subgenera, as suggested by Karaman, 1959, Ruffo (1970), Stock, 1976, and Ruffo & Vigna Taglianti (1989). However, even though Vonk & Schram’s manuscript represents the most complex and exhaustive work available on Ingolfiellidae phylogeny and biogeography, we think some of the results still need further discussion, which, however, is not the aim of the present paper. The CI value of 0.3511 and a HI value of 0.6489 reported in the cladistic analysis of Vonk & Schram (2003) do not represent optimal values to provide an interpretation of the results. Moreover, “only 13 of the 27 species of the genus Ingolfiella have both sexes well known… 11 are known only for females and three are known only from males” ( Senna & Serejo, 2005), and therefore several characters related to sexual dimorphism were not taken into account by Vonk & Schram ( op. cit.) for cladistic analysis. Several taxa are poorly described when compared with the recent taxonomic descriptions. In conclusion, our opinion is that the systematic of the genus is still in state of flux and therefore it may be too early to institute new genera and/or subgenera of Ingolfiella . In agreement with Vonk & Schram ( op. cit.) “as more taxa are added to the genus the cladogram will acquire some additional structure”, and so, it will be possible to review the systematic of the genus. In the meantime the species-groups sensu Ruffo (1970) and Ruffo & Vigna Taglianti (1989) may still be of practical taxonomic use. Furthermore, more detailed biogeographic data are required to reconstruct the history of the actual distribution of Ingolfiellidae . For the genus Ingolfiella there is still a need for alpha-taxonomy, using the “traditional” characters but also other, informative characters which were often ignored even in recent description, such as the structure of transformed setae and spines in mouthparts, pereopods, and uropods. Vonk & Schram ( op. cit.) presented a long and interesting discussion of the distribution of the genus Ingolfiella , suggesting that in this taxon, as in other peracarids, the marine interstitial species, which typically have reduced body size, are the most recent ones. They derived from freshwater cave ancestors in a three-steps process leading first to freshwater interstitial species of small size from which (about 90 my) would have originated first brackish, and then marine taxa living in interstitial and deep benthic habitats. There are other alternative hypotheses to this scenario, which are discussed in Vonk & Scram ( op. cit.). Finally, it is still not possible to choose one of the several hypotheses proposed because from one hand the Ingolfiellidae phylogeny is not completely clear, and on the other hand, the number of known species and data on the genus geonemy are still insufficient.
Sampling sites and remarks on the ecology. The two sampling sites have similar substrate, represented by medium-coarse carbonatic sand, composed by shell and coral fragments. The holes from where the specimens were collected were dug on the beach during low tide, 2–3 m from the shoreline. In Marinduque, the pH of interstitial water was 8.1 and the temperature 28.4ºC, while in Mindoro, the pH was 7.8 and the temperature, 24.7ºC. Ingolfiella alba sp. nov. was collected together with isopods of the genus Microcerberus Karaman, 1933 and several harpacticoid taxa. Palpigrads probably of the genus Leptokoenenia Condè 1965 were collected in Marinduque stations. One of these palpigrads was holding a half-eaten male of I. alba sp. nov on the chelicera. This information might be relevant because there are no literature data reporting predation on Ingolfiellidae , and data on the autoecology of Leptokoenenia , as well as for all palpigrads, are very limited ( Condé, 1996). The interstitial habitat is in fact characterized by the scarcity of specialized predators, thus the importance of reporting any new information on such interactions among interstitial organisms.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ingolfiella alba
| Iannilli, Valentina, Berera, Raffaella & Cottarelli, Vezio 2008 |
Metaingolfiella mirabilis
| Ruffo 1969 |