
Imparfinis usmai, Ortega-Lara & Milani & DoNascimiento & Villa-Navarro & Maldonado-Ocampo, 2011
|
publication ID |
https://doi.org/10.1590/S1679-62252011000400009 |
|
DOI |
https://doi.org/10.5281/zenodo.5072755 |
|
persistent identifier |
https://treatment.plazi.org/id/03C92A47-FFAB-480E-CECF-E2DAEC29F805 |
|
treatment provided by |
Carolina |
|
scientific name |
Imparfinis usmai |
| status |
sp. nov. |
Imparfinis usmai View in CoL , new species
Fig. 3 View Fig
Holotype. IMCN 4812 , 83.4 mm SL, Colombia, Departamento del Tolima, Municipio Coello, vereda Potrerillo , quebrada Potrerilla , tributary of río Coello , río Magdalena basin, 04°16’57.9”N 75°01’53.8”W, 13 Jan 2007, A. Ortega-Lara & F. Villa-Navarro. GoogleMaps
Paratypes: Colombia: río Cauca basin: Departamento del Cauca: IMCN 168 , 9 , 62.6-88.4 mm SL, Municipio Suarez, río Cauca, km 21 in the road to Suárez , 03°25’N 76°40’W, 10 Sep 2002,A. Ortega- Lara. IMCN 2301 , 7 , 48.1-84.9 mm SL, GoogleMaps Municipio Santander de Quilichao, río Quinamayó , road to La Balsa , 03°25’N 76°35’W, 26 Jun 2002, A. Ortega-Lara. IMCN 2439 , 7 , 37.5-75.6 mm SL, GoogleMaps Municipio Santander de Quilichao, mouth of río Quinamayó in río Cauca, 03°25’N 76°38’W, 23 Jul 2002, A. Ortega-Lara. IMCN 3842 , 10 , 40.6-73.3 mm SL, GoogleMaps Municipio Caloto , río Japio , at Hacienda Japio , road to Miranda , 03°01’N 76°24’W, 7 Oct 2004, A. Ortega- Lara. IMCN 3914 , 8 , 47.5-89.3 mm SL (1 CS, 82.5 mm SL), GoogleMaps Municipio Guachené , río Palo , at the bridge of Guachené , 03°07’N 76°22’W, 17 Jun 2002, A. Ortega-Lara. Departamento del Valle del Cauca: CZUT-IC 1158 , 2 , 51.3-55.3 mm SL, río Cauca, 25 Jun 2002, A. Ortega-Lara. IMCN 1164 , 2 , 23.0- 79.5 mm SL, GoogleMaps Municipio Cartago , río La Vieja , feeder of irrigation district, 04°45’39.4”N 75°53’63.2”W, 990 m asl, 18 Jul 2002, S. Usma, L. Meza, B. Arias, W. Arias & C. Araque. IMCN 1166 , l, 37.2 mm SL, same locality and collectors as IMCN 1164 , 24 Jun 2002. IMCN 1179 , 5 , 48.8- 92.6 mm SL, MBUCV-V-30943, 2, 71.3-89.1 mm SL (1 CS, 89.1 mm SL), MBUCV-V-35640, 88.7 mm SL, same locality and collectors as IMCN 1164 , 21 Aug 2002. IMCN 1176 , 7 , 53.9-86.8 mm SL (2 CS, 53.9-85.8 mm SL), MBUCV-V-33734, 3, 61.1-73.7 mm SL (1 CS, 61.1 mm SL), GoogleMaps Municipio Cartago, río La Vieja, sector Piedras de Moler, road to Alcalá, 04°42’32.6”N 75°51’36”W, 1100 m asl, 18 Jul 2002, same collectors as IMCN 1164 . IMCN 1178 , 3 , 48.5 -75.0 mm SL, same locality as IMCN 1176, 24 Jun 2002, S. Usma, L. Meza, B. Arias, W. Arias & C. Araque. IMCN 1180 , 1 , 72.8 mm SL, GoogleMaps Municipio Cartago, río La Vieja, at the airport, 4°47’18.9”N 75°55’71.0”W, 980 m asl, 18 Jul 2002, same collectors as IMCN 1164 . IMCN 3277 , 1 CS, 76.3 mm SL, GoogleMaps Municipio Jamundí, río Cauca, sector El Hormiguero, 02°42’25”N 76°41’37”W, Mar 2003, A. Ortega-Lara. IMCN 3278 , 1 CS, 64.5 mm SL, GoogleMaps Municipio Ansermanuevo, río Chanco , sector Hacienda Arauca, 04°47’50”N 75°59’42”W, 920 m asl, 18 Mar 2002, A. Ortega-Lara. Departamento del Quindío: IMCN 1170 , 3 , 67.5-83.2 mm SL (1 CS, 67.5 mm SL), Municipio Armenia, río Quindío, Tarapacá, 04°23’74”N 75°45’94.9”W, 1130 m asl, 22 Aug 2002, S. Usma, L. Meza, B. Arias W. Arias & C. Araque. Departamento de Risaralda: IAvH-P 7160, 4, 55.0- 70.7 mm SL, IMCN 3373 , 8 , 33.3-85.6 mm SL, GoogleMaps Municipio La Virginia , río Risaralda, downstream of mouth of río Mapa , 04°55’N 75°51’W, 24 Feb 2005, A. Ortega-Lara. Río Magdalena basin: Departamento del Tolima: CZUT-IC 91 , 1 , 84.9 mm SL, GoogleMaps Municipio Coello, vereda Potrerillo, quebrada Potrerilla, tributary of río Coello , 04º16’57.9”N 75º01’53.8”W, 21 Mar 2003, A. Ortega-Lara, L. García-Melo, P. Zúñiga-Upegui, N. Briñez- Vasquez & F. Villa-Navarro. CZUT-IC 214 , 4 , 63.3-88.9 mm SL, same locality and collectors as CZUT-IC 91 , 2 May 2003. CZUT- IC 98, 15, 60.9-100.8 mm SL (1 CS, 60.9 mm SL), GoogleMaps Municipio Coello, quebrada Gualanday, 04°18’17.5”N 75°02’01”W, 9 May 2003, A. Ortega-Lara, L. García-Melo, P. Zúñiga-Upegui, N. Briñez- Vasquez & F. Villa-Navarro. CZUT-IC 184 , 10 , 70.8-90.5 mm SL, GoogleMaps Municipio Coello, río Coello , 50 m downriver from feeder of irrigation district USOCOELLO, Inspección de Gualanday , 04°16’50.3”N 75°01’50.7”W, 28 Mar 2003, A. Ortega-Lara, L. García-Melo, P. Zúñiga-Upegui, N. Briñez-Vasquez & F. Villa- Navarro. CZUT-IC 637 , 3 , 65.7-89.2 mm SL, same locality and collectors as CZUT-IC 184 , 20 Sep 2003. CZUT-IC 792 , 4 , 65.9- 96.3 mm SL, same locality and collectors as CZUT-IC 184 , 27 Jul 2003. CZUT-IC 229 , 3 , 82.8-92.7 mm SL, same locality as CZUT- IC 184, 2 May 2003, GoogleMaps A. Ortega-Lara, L. García-Melo, P. Zúñiga- Upegui, N. Briñez-Vasquez & F. Villa-Navarro. CZUT-IC 874 , 30 , 57.0- 85.7 mm SL, río Tetuán , 03°45’N 75°20’W, 5 Oct 2003, A. Ortega-Lara, L. García-Melo, P. Zúñiga-Upegui, N. Briñez-Vasquez & F. Villa-Navarro. CZUT-IC 892 , 8 , 20.9-90.8 mm SL, same locality as CZUT-IC 874, 6 Jun 2003, GoogleMaps A. Ortega-Lara, L. García-Melo, P. Zúñiga-Upegui, N. Briñez-Vasquez & F. Villa-Navarro. CZUT-IC 1053 , 1 , 70.8 mm SL, río Anchique , 03º34’34.8”N 75º07’13”W, 21 Mar 2004, A. Ortega-Lara, L. García-Melo, P. Zúñiga-Upegui, N. Briñez-Vasquez & F. Villa-Navarro. CZUT-IC 1155 , 5 , 28.8-51.3 mm SL, quebrada Silletero, tributary of río Cauca, 10 Feb 2004, A. Ortega-Lara. CZUT-IC 1158 , 2 , 50.6 -56.0 mm SL, río Cauca, 21 Jun 2002, GoogleMaps A. Ortega-Lara. CZUT-IC 1345 , 11 , 31.5-44.2 mm SL, mouth of río Amoya , 03º40’23.7”N 75º23’24.5”W, 8 Feb 2005, L. García-Melo, N. Briñez-Vasquez, D. Castro-Roa, M. Herrada-Yara & F. Villa-Navarro. CZUT-IC 1379 , 2 , 53.3-57.2 mm SL, GoogleMaps Municipio Chaparral, cuevas de Tuluni, vereda Tuluni, 03º38’52.2”N 75º27’24.5”W, 9 Jan 2005, L. García-Melo, N. Briñez-Vasquez, D. Castro-Roa, M. Herrada-Yara & F. Villa-Navarro. CZUT-IC 1419 , 2 , 41.2-49.5 mm SL, CZUT-IC 1431 , 10 , 45.8-76.5 mm SL, GoogleMaps Municipio Chaparral, quebrada Tuluni, vereda Tuluni, 03º39’08.9”N 75º29’05”W, 10 Feb 2005, L. García-Melo, N. Briñez-Vasquez, D. Castro-Roa, M. Herrada-Yara & F. Villa-Navarro. IMCN 3847 , 7 , 25.1-96.4 mm SL, MBUCV-V-33736, 4, 60.3-89.7 mm SL (1 CS, 60.3 mm SL), collected with the holotype GoogleMaps .
Non-type material. Colombia: río Patía basin: Departamento del Cauca : FMNH 58131 About FMNH , 2 About FMNH , 61.2-68.9 mm SL, río Telembí, downstream from Barbacoas, lower basin tributary of río Patía . Río Cauca basin: Departamento del Valle del Cauca : FMNH 58129 About FMNH , 3 About FMNH , 67.2-78.7 mm SL, río Cauca at Cali . FMNH 58132 About FMNH , 8 About FMNH , 32.3-40.8 mm SL, Cali , 1010 m asl . IMCN 2475 , 1 , 35.2 mm SL, Municipio Cartago, río La Vieja, sector Piedras de Moler , road to Alcalá , 04°42’32.6”N 75°51’36”W, 1100 m asl GoogleMaps . IMCN 2635 , 4 , 61.2-69.6 mm SL, MunicipioAnsermanuevo, río Cañaveral , to Ansermanuevo, La Virginia . MCZ 35873, 2 About MCZ , 30.6-36.8 mm SL, upper río Cauca. Departamento del Quindío : IMCN 2394 , 2 , 38.3-59.8 mm SL, Municipio Armenia, río Quindío, Tarapacá , 04°23’74”N 45°45’94.9”W, 1130 m asl .
Diagnosis. This species is distinguished from congeneric trans-Andean species by having the first ray of dorsal fin longest and slightly projected or not beyond dorsal-fin margin, length of first dorsal-fin ray: 19.1-23.2% in SL ( vs. slightly shorter than second ray in I. lineatus , 14.0-17.6%; I. spurrellii , 16.0-17.6%; and I. timana , 16.3-21.0%; or extended as a conspicuous and long filament beyond fin margin in I. nemacheir , 29.2-33.1%); and by the intermediate long upper caudal-fin lobe (24.4-39.2% in SL vs. 22.7-26.0% in I. lineatus ; 38.2-52.1% in I. nemacheir ; 22.8-26.5% in I. spurrellii ; 21.9- 28.0% in I. timana ). Imparfinis usmai differs from all trans- Andean species, except I. nemacheir , by having a longer maxillary barbel (40.3-63.8% in SL vs. 24.0-32.6% in I. lineatus ; 32.1-35.2% in I spurrellii ; 32.3-45.0% in I. timana ), and by the number of vertebrae (39-40 vs. 42 in I. lineatus ; 45 in I. spurrellii ; 42-43 in I. timana ). Specimens of Imparfinis usmai exceeding 30 mm SL, are readily distinguished from all trans- Andean species, except I. timana , by having an upper caudalfin lobe pointed and longer than lower lobe, and a lower lobe rounded, with both branches of lowermost branched ray approximately equal in length ( vs. both lobes rounded and approximately symmetrical in I. lineatus and I. spurrellii ; both lobes pointed, with lower branch of lowermost branched ray longer than upper branch, and extended as a filament in I. nemacheir ). Imparfinis usmai can be recognized from I. lineatus and I. nemacheir by the longer adipose-fin (21.1- 27.0% in SL vs. 17.2-18.6% in I. lineatus ; 19.0-21.8% in I. nemacheir ). Imparfinis usmai is further distinguished from I. lineatus and I. timana by its shorter caudal peduncle depth (5.6-7.8% in SL vs. 8.1-8.4% in I. lineatus ; 7.1-8.7% in I. timana ). Imparfinis usmai is further recognized from I. spurrellii and I. timana by having 7-8 gill rakers on the first ceratobranchial ( vs. 10-11 in I. spurrellii ; 5-6 in I. timana ). Imparfinis usmai can also be distinguished from I. spurrellii by its greater interorbital width (27.8-36.3% in HL vs. 23.6-24.3% in I. spurrellii ). Specimens of Imparfinis usmai exceeding 30 mm SL differs from all cis-Andean congeners, except I. microps , I. mirini , I. mishky , and I. piperatus , by having a forked caudal fin, with upper lobe longer and pointed, and lower lobe rounded ( vs. both caudal-fin lobes pointed). Imparfinis usmai is further distinguished from I. borodini and I. hollandi by having longer maxillary barbels (surpassing pelvic-fin base vs. not exceeding pectoral-fin distal margin), fewer vertebrae (39-40 vs. 49-54), insertion of first dorsal-fin pterygiophore posterior to neural spine of vertebra 7 ( vs. vertebra 12-13), pelvic-fin origin at or slightly posterior to mid-distance of dorsal-fin base ( vs. at vertical or slightly displaced from dorsalfin origin), fewer total anal-fin rays (12-13 vs. 14-15), insertion of first anal-fin pterygiophore posterior to hemal spine of vertebra 23 ( vs. vertebra 29-31), caudal fin deeply forked ( vs. caudal fin obliquely truncated with upper end prolonged), and more branched caudal-fin rays (7+8 vs. 6+5-6); additionally I. usmai is distinguished from I. borodini by having a shorter adipose fin (3.70-4.74 times in SL vs. 2.67 times); and from I. hollandi by having fewer pleural ribs (8-9 vs. 11), and adipose fin free from caudal fin ( vs. continuous). Imparfinis usmai is recognized from I. pseudonemacheir and I. stictonotus by having shorter maxillary barbels (not reaching adipose-fin origin vs. extending to first half of adipose-fin base or beyond), more vertebrae (39-40 vs. 34-38), and sides of body without distinct markings ( vs. four irregular dark patches); additionally I. usmai is distinguished from I. pseudonemacheir by having a larger size (maximum SL: 100.8 mm vs. 48.1 mm), fewer gill rakers on first gill arch (8-10 vs. 11-14), distal margin of adpressed anal fin anterior to adipose-fin posterior end ( vs. posterior), upper caudal-fin lobe longer than lower lobe ( vs. both caudal-fin lobes equal in length); and from I. stictonotus by having more branched anal-fin rays (7-8 vs. 6), both caudalfin lobes similarly pigmented ( vs. lower lobe darker than upper lobe). Imparfinis usmai can be differentiated from remaining cis-Andean congeners as follows: from I. cochabambae by having fewer gill rakers on first gill arch (8-10 vs. 15), and fewer vertebrae (39-40 vs. 41); from I. guttatus by having fewer gill rakers on first gill arch (8-10 vs. 14), longer adipose fin (3.70-4.74 times vs. ca. 5.65 times, obtained from holotype picture), and both caudal-fin lobes similarly pigmented ( vs. lower caudal-fin lobe darker); from I. hasemani by having longer maxillary barbels (surpassing pelvic-fin base vs. not exceeding pectoral-fin distal margin), and fewer pleural ribs (8-9 vs. 10); from I. longicauda by having a longer head (4.08- 5.15 times in SL vs. 6 times or more), and insertion of first dorsal-fin pterygiophore posterior to neural spine of vertebra 7 ( vs. vertebra 8); from I. microps by having longer maxillary barbels (surpassing pelvic-fin base vs. not extending beyond posterior margin of branchiostegal membrane), fewer branchiostegal rays (6-7 vs. 9), more gill rakers on first gill arch (8-10 vs. 4-5), fewer vertebrae (39-40 vs. 46-47), more branched pectoral-fin rays (9 vs. 7), more posterior pelvic-fin origin (at or slightly posterior to mid-distance of dorsal-fin base vs. anterior to dorsal-fin origin), insertion of first dorsalfin pterygiophore posterior to neural spine of vertebra 7 ( vs. vertebra 15), fewer branched anal-fin rays (7-8 vs. 9), anal-fin origin posterior to adipose-fin origin ( vs. anterior), insertion of first anal-fin pterygiophore posterior to hemal spine of vertebra 23 ( vs. vertebra 26), and more branched caudal-fin rays (7+8 vs. 6+6); from I. minutus by having longer maxillary barbels (surpassing pelvic-fin base vs. not exceeding distal tip of adpressed pectoral fin), fewer vertebrae (39-40 vs. 41), and insertion of first dorsal-fin pterygiophore posterior to neural spine of vertebra 7 ( vs. vertebra 8-9); from I. mirini by having longer maxillary barbels (surpassing pelvic-fin base vs. scarcely beyond distal tip of adpressed pectoral fin), adipose-fin posterior end extending more posteriorly than distal margin of adpressed anal fin ( vs. approximately at same level), and upper caudal-fin lobe longer than lower lobe ( vs. both caudal-fin lobes equal in length); from I. mishky by having fewer branched rays on lower lobe of caudal fin (8 vs. 9), shorter anal-fin base (10.8-14.5% in SL vs. 13.1-16.3%), longer caudal peduncle (19.8-25.3% in SL vs. 17.7-20.1%), and greater interorbital width (27.8-36.3% in SL vs. 18.5-23.1%); from I. peruanus by having a shorter predorsal distance (2.75-3.37 times in SL vs. 2.6 times), insertion of first dorsal-fin pterygiophore posterior to neural spine of vertebra 7 ( vs. vertebra 9), and insertion of first anal-fin pterygiophore posterior to hemal spine of vertebra 23 ( vs. vertebra 25); from I. pijpersi by having longer maxillary barbels (surpassing pelvic-fin base vs. not exceeding distal tip of adpressed pectoral fin), longer caudal peduncle (19.8-25.3% in SL vs. 16.1%), shorter caudal peduncle depth (5.6-7.8% in SL vs. 11.0%), and longer adipose fin (21.1-27.0% in SL vs. 13.1%); from I. piperatus by having longer maxillary barbels (surpassing pelvic-fin base vs. reaching distal tip of adpressed pectoral fin), more vertebrae (39-40 vs. 37), and insertion of first anal-fin pterygiophore posterior to hemal spine of vertebra 23 ( vs. vertebra 20); from I. pristos by having a larger size (maximum SL: 100.8 mm vs. 38.0 mm, value from Bockmann & Guazelli, 2003), shorter head (4.08-5.15 times in SL vs. 3.4- 3.7), longer maxillary barbels (surpassing pelvic-fin base vs. not exceeding distal tip of adpressed pectoral fin), more gill rakers on first gill arch (8-10 vs. 4-6), more vertebrae (39-40 vs. 33-35), more pleural ribs (8-9 vs. 6), insertion of first dorsal-fin pterygiophore posterior to neural spine of vertebra 7 ( vs. vertebra 6), longer adipose fin (3.70-4.74 times in SL vs. 5.0- 5.6 times), more branched anal-fin rays (7-8 vs. 5-6), insertion of first anal-fin pterygiophore posterior to hemal spine of vertebra 23 ( vs. vertebra 17-18), upper caudal-fin lobe longer than lower lobe ( vs. both caudal-fin lobes equal in length), and more branched rays on caudal fin (7+8 vs. 6+7); and from I. schubarti by having longer maxillary barbels (surpassing pelvic-fin base vs. reaching or scarcely beyond distal tip of adpressed pectoral fin), fewer gill rakers on first gill arch (8-10 vs. 11-16), longer adipose fin base (21.1-27.0% in SL vs. 18.7- 22.6%), and shorter preanal length (63.1-71.4% in SL vs. 71.0- 74.7%).
Description. Morphometric data given in Table 1 View Table 1 . Small heptapterid catfish (largest specimen 100.8 mm SL), with elongated body, triangular in cross-section at dorsal-fin origin, progressively more laterally compressed to tail region. Dorsal profile slightly convex from snout tip to dorsal-fin origin, slightly concave just posterior to dorsal-fin base to adiposefin origin, straight and descending along adipose-fin base, then ascending along caudal peduncle. Ventral profile of head straight, slightly convex along abdomen, then straight to analfin origin and slightly descending along caudal peduncle.
Head conical and depressed, dorsally covered by thin skin. Snout short and broadly rounded. Mouth subterminal. Premaxillary teeth conical and pointed arranged in rectangular band with 4-5 irregular rows. Dentary with 4-5 irregular rows of teeth. Barbels dorsoventrally flattened. Maxillary barbels surpassing pelvic-fin base. Anterior portion of maxillary barbel in shallow groove, extending below eye. Bases of outer and inner mental barbels aligned. Outer mental barbels surpass pectoral-fin base. Inner mental barbels reach pectoral-fin origin. Eye dorsolateral in position. Orbital margin not free, but delimited by shallow groove, more conspicuous along dorsal rim. Anterior naris tubular. Posterior naris triangular, slightly closer to anterior ocular margin than to anterior naris, bordered by low fleshy margin, restricted to anterolateral rim. Nares disposed in trapezoidal arrangement. Anterior internarial width slightly shorter (mean % in HL 17.5) than posterior internarial width (mean % in HL 21.6). Branchiostegal membrane free, supported by 6-7 rays and joined to isthmus only at anteriormost point. Gill rakers on first gill arch 8-10, 7-8 arranged on anterior margin of ceratobranchial, one on cartilage between ceratobranchial and epibranchial and 0-1 on epibranchial.
Lateral line canal complete, extending to basal portion of interradial membrane of middle caudal-fin rays. Supraorbital pore S1 medially adjacent to anterior naris. S2+I2 between anterior and posterior nares, slightly closer to posterior naris, at end of posteriorly directed membranous tubule originating from commissure connecting supraorbital and infraorbital canals, closer to supraorbital canal. S3 just at posterior rim of posterior naris and S4 located posterior to posterior naris and at level of anterior eye margin. Both S3 and S4 originating from anterior and posterior ends of bifurcated lateral membranous branch with T-shape. S8 (corresponding to parietal branch), posteromedial to eye. Parietal branch running posteriorly on frontal bone, and ending close to articular suture with parieto-supraoccipital bone. Infraorbital pore I1 laterally adjacent to anterior naris, just between naris and maxillary barbel base. I3 posterior to maxillary barbel base. I4 at vertical through anterior eye margin. I5 at vertical through posterior eye margin. I6 posterior to eye at end of short ventroposterior membranous tubule. Preoperculomandibular canal with 11 pores. Dentary with seven pores. Submental pores (PM1) paired and last mandibular pore (PM7) at level of articulation between anguloarticular and quadrate bones. Preopercle with three pores. Anterior pore (PM8) originating between subpreopercle and preopercle. Middle pore (PM9) originating from membranous tubule passing above interopercle and posterior pore (PM10) from membranous tubule passing above ventral portion of opercle. Last preopercular pore (PO1+PM11) at end of membranous branch, at level of posterior process of hyomandibula. PO2 corresponding to pterotic branch, located dorsal to dorsoposterior corner of opercular margin. Axillary branch (LL1) ventral, running posterior to extrascapular. Accessory branch (PO3) dorsal to lateral line canal, ending approximately at axilar pore level.
Precaudal vertebrae 11-13 and caudal vertebrae 27-29, totaling 39-40 vertebrae. First hemal spine on vertebra 15-16. Pleural ribs eight or nine. Anus approximately at mid-length of pelvic fin, closer to pelvic-fin base than anal-fin origin. Urogenital papilla separated from anus by distance approximately equivalent to length of papilla.
Pectoral fin i,9. Basal portion of simple ray ossified, distal portion soft and segmented. Pectoral fin triangular, first ray sometimes slightly projected beyond fin margin. Distal margin of pectoral fin slightly convex. Pelvic fin i,5. First pelvic-fin ray thick and shortest, first and second branched rays longest. Pelvic-fin origin at or slightly posterior to middistance of dorsal-fin base. Dorsal fin lacking spinelet ( i.e. first lepidotrichium), with one simple (second lepidotrichium), and six branched rays. Dorsal fin triangular, first ray longest, sometimes slightly projected beyond fin margin to less than 10% length of second ray. Supporting fin elements represented by seven proximal and six distal radials. Last two branched rays articulating separately with last two pterygiophores. First proximal radial inserted posterior to neural spine of vertebra 7 and last proximal radial inserted anterior to neural spine of vertebra 13-14. Adipose fin low, its maximum height at anterior third, and longer than anal fin, with free posterior lobe. Adipose-fin origin anterior to anal-fin origin.Anal fin with 2-3 procurrent rays, associated with first proximal radial, 2-3 simple rays and 7-8 branched rays (total of ten principal rays). Anal-fin distal margin rounded. Anal fin supported by 9-10 proximal and 8-9 distal radials. First proximal radial posterior to hemal spine of vertebra 23 and last proximal radial anterior to hemal spine of vertebra 28-29. Caudal fin deeply forked with i,7+8,i principal rays. Upper caudal-fin lobe pointed and longer than lower lobe in adult specimens and juveniles longer than 30 mm SL; both lobes pointed but not prolonged as filaments, and about same size, in juvenile specimens shorter than 30 mm SL. Lower caudal-fin lobe rounded in specimens exceeding 30 mm SL, with both branches of lowermost branched ray approximately equal in length. Dorsal procurrent caudal-fin rays 14-17, located posterior to vertebrae PU 5 -PU 6. Last two rays segmented. Ventral procurrent caudal-fin rays 13-14, located posterior to vertebrae PU 6 -PU 7. Last three rays segmented. Caudal skeleton PH, 1+2, 3+4, 5 (MBUCV-V-30943 CS specimen with hypurals 3+4 partially fused with 5 at base). Long epural present.
Coloration. Live specimens as shown in Fig. 4 View Fig , dorsal and lateral surface of body purplish brown, abdominal surface cream. Dorsal and lateral surface of head and dorsal surface of predorsal region greenish yellow. Dorsal surface of barbels brown, and ventral surface of mental barbels white. Gold iridescent blotch on opercular region. Four dark brown saddles on dorsum, first saddle crossing predorsal region, second saddle just in front of dorsal-fin origin, third saddle on posterior half of dorsal-fin base, and last saddle between dorsal and adipose fins. Humeral region with black diffuse blotch continuous posteriorly with midlateral diffuse band. Fin rays of dorsal, pectoral, pelvic, and caudal fins yellowish brown. Interradial membrane of dorsal fin hyaline and membrane of pectoral fin light yellow.Adipose fin light yellow, with dark chromatophores disperse on fin base. In alcohol, body brownish on dorsal and lateral surfaces and cream on ventral surface. Conspicuous dark mid-lateral stripe, extending from posterior margin of opercle to caudal-fin origin. Dorsal surface of maxillary barbels pigmented. Dorsal surface of mental barbels with sparse chromatophores. Parieto-supraoccipital region densely pigmented. Opercle with dark blotch, some chromatophores grouped below eye, along region corresponding to preopercle. Rays of pectoral, dorsal, anal, and caudal fins darkly pigmented. Interradial membranes hyaline. Mid-portion of pectoral and pelvic-fin rays with chromatophores scattered and distal margin hyaline. Adipose fin with chromatophores at basal portion and distal margin hyaline.
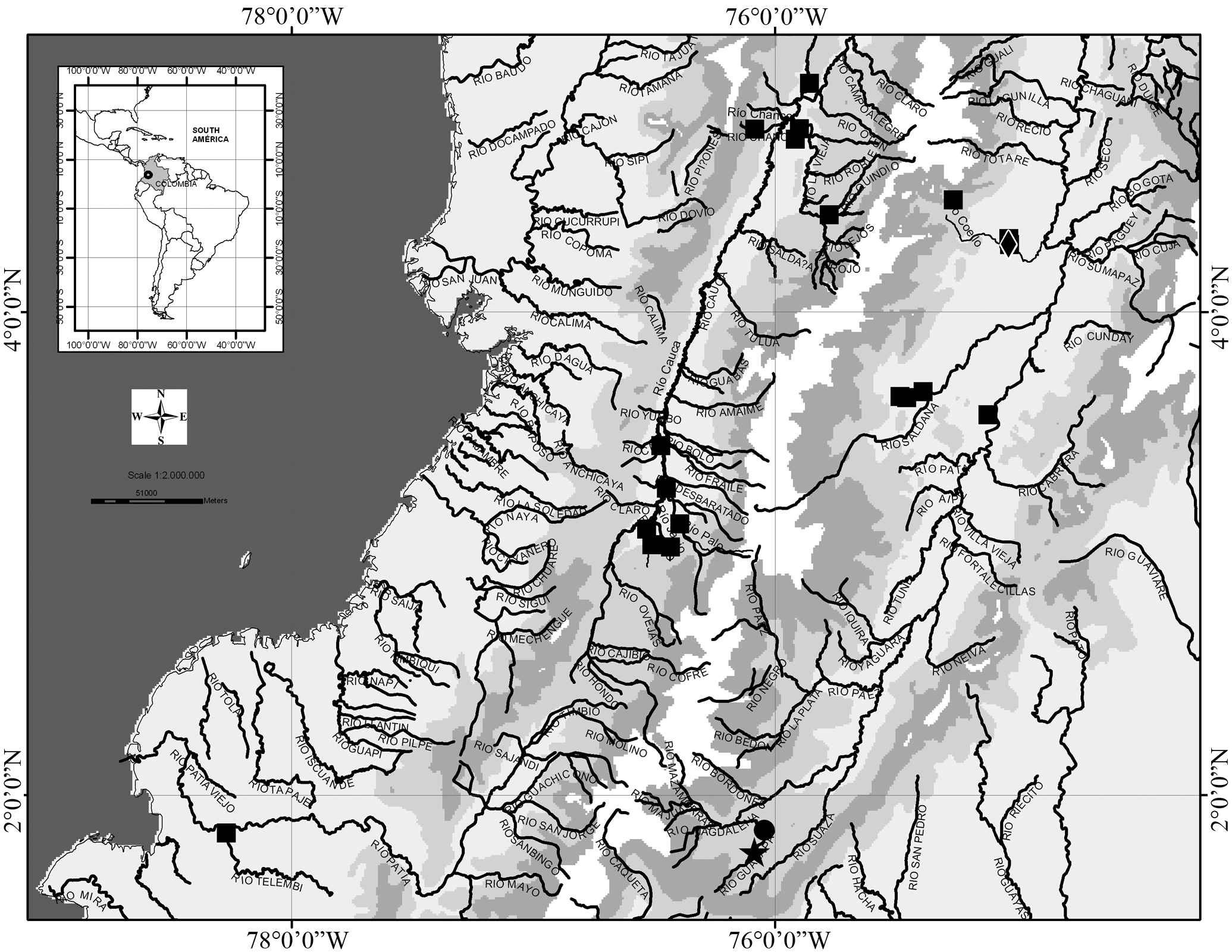
Distribution. This species is broadly distributed along the upper basin of río Cauca and río Magdalena, and in the lower río Patía basin on the Pacific slope of western cordillera of Colombia ( Fig. 2 View Fig ). Imparfinis usmai is sympatric with I. nemacheir in the río Magdalena.
Etymology. Dedicated to Saulo Usma for his contributions to the ichthyological collection building in the Cauca valley, and for promoting an active interchange between Colombian and Venezuelan ichthyologists, since his MSc studies in 2000 at Universidad Nacional Experimental de los Llanos Ezequiel Zamora (Guanare, Venezuela).
Remarks. Proper taxonomic recognition of Imparfinis usmai has an especially confusing record in the ichthyological literature, even in strictly taxonomic works. Eigenmann (1922), while redescribed Imparfinis nemacheir , listed a series of specimens coming from the ríos Patía and Cauca, as reference material for this species. Some of these specimens (FMNH 58129, 58131-32), formerly catalogued at the Carnegie Museum, were reexamined, and we found that these population samples are assignable to I. usmai . This species has also been misidentified as I. nemacheir in Ortega-Lara et al. (2000) (photograph on p. 41, where the scarcely projected first dorsal-fin ray beyond fin membrane and shorter maxillary barbel are evident when compared to the actual I. nemacheir ), and in Maldonado-Ocampo et al. (2005) (illustration of Fig. 154, p. 289, showing the characteristic configuration of the caudal fin of I. usmai , with upper lobe longer and pointed and lower lobe rounded; along with most of the reference material therein listed and reidentified in the present work: see CZUT-IC and IMCN lots of paratypes and non-type material of I. usmai ). Even more surprising are the identifications of pictured specimens of I. usmai as Rhamdia sebae (Cuvier, 1829) (photograph in p. 70) in Galvis et al. (1997), and as Cetopsorhamdia nasus Eigenmann & Fisher, 1916 ( Fig. 2 View Fig , p. 128) in Ruiz-C. & Román-Valencia (2006). From our revision we can assume that some of the distribution records attributed to Imparfinis nemacheir in ríos Magdalena and Cauca basins, in fact correspond to Imparfinis usmai . This last species ( Fig. 5a View Fig ) being easily differentiated from the sympatric I. nemacheir ( Fig. 5 View Fig b-d) by a series of apparent external features briefly mentioned above, as well as those provided in the identification key, which are exhaustively compared in I. usmai diagnosis.
Those lots listed as non-type material correspond to very old specimens (collection dates: 1912-1913), that do not exhibit all diagnostic characters for the species in every single individual, perfectly understandable given the long history of preservation and manipulation ( e.g. broken fins); or more recently collected specimens that are not ideally preserved, with distorted body, broken barbels or fins.



| CS |
Musee des Dinosaures d'Esperaza (Aude) |
| MCZ |
Museum of Comparative Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |