
Ilyocypris tibeta, Peng & Zhai & Smith & Wang & Guo & Zhu, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4942.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:EB79F747-805C-46AB-BBC1-191191B860A2 |
|
DOI |
https://doi.org/10.5281/zenodo.4644595 |
|
persistent identifier |
https://treatment.plazi.org/id/ED6E87E9-FFEB-FFAB-FF66-F92AFEE56CC1 |
|
treatment provided by |
Plazi |
|
scientific name |
Ilyocypris tibeta |
| status |
sp. nov. |
Ilyocypris tibeta n. sp.
( Figs 18–22 View FIGURE 18 View FIGURE 19 View FIGURE 20 View FIGURE 21 View FIGURE 22 )
? 2009a Ilyocypris cf. mongolica Martens, 1991 —Wrozyna et al.: 8, 12, 14–17, plate 1: 1–8, figs 6–8.
? 2009b Ilyocypris cf. mongolica Martens, 1991 —Wrozyna et al.: 672, 673, plate 3: 7–11.
? 2013 Ilyocypris cf. bradyi Sars, 1890 —Zhang et al.: 40–43, 45, figs 2, 4, 6 (6–10).
? 2015a Ilyocypris bradyi — Song et al.: 587, 589–592, figs 1 (1&2), 3, 4 table 1.
? 2016 Ilyocypris sp.— Akita et al.: 27–30, 35, 37, 38, Figs 3 View FIGURE 3 (16–18), 7, 9, 10, 12, tables 4, 5, 6.
Type locality. A shallow pond in Rutog , Ngari, Tibet, China (32.93723ºN, 79.80317ºE, altitude 4390 m) ( OS11-14 , Table 1 View TABLE 1 ) GoogleMaps .
Type material. Holotype: a dissected male ( WOC39 ) from type locality ( Table 1 View TABLE 1 ) . Allotype: a dissected female ( WOC38 ) from type locality . Paratypes: two dissected females ( WOC40 , WOC45) and two dissected males ( WOC36 , WOC37) ( Table 1 View TABLE 1 ). Carapace of WOC40 damaged during dissection .
Other material examined. One dissected, probably male specimen ( WOC 35) with soft parts deteriorated (only Md coxa remained).
Derivation of name. From name of the province where the specimens were collected.
Dimensions. Male (LV, n = 3, WOC35 not included) length 0.77–0.89 mm, H/L ratio 0.52–0.54. Female (LV, n = 3) length 0.83–0.98 mm, H/L ratio 0.51–0.52.
Diagnosis. Medium-sized Ilyocypris . Shell surface covered with small, rounded pits. In interior view, LV with numerous sparsely distributed marginal ripplets in distal area of posterior inner calcified lamella. A2 natatory setae extending to tips of terminal claws. Male L5 palps sub-equal, with sub-apical part of fingers slightly inflated. L6 penultimate segment divided. L7 penultimate segment with two setae (f and g) and apically bearing ca. four unequal length, hook-like structures. Inner lobe (b) of Hp elongate and slender, distally curved towards middle lobe. Middle lobe (h) with relatively narrow neck distally widening to sub-triangular distal part, distal inner corner sharply angled, outer edge angled to rounded. Outer lobe (a) short, distally evenly rounded and partly overlapping middle lobe. Copulatory process (st) slender, with pointed end. ZO with 16–17 internal rosettes of spines.
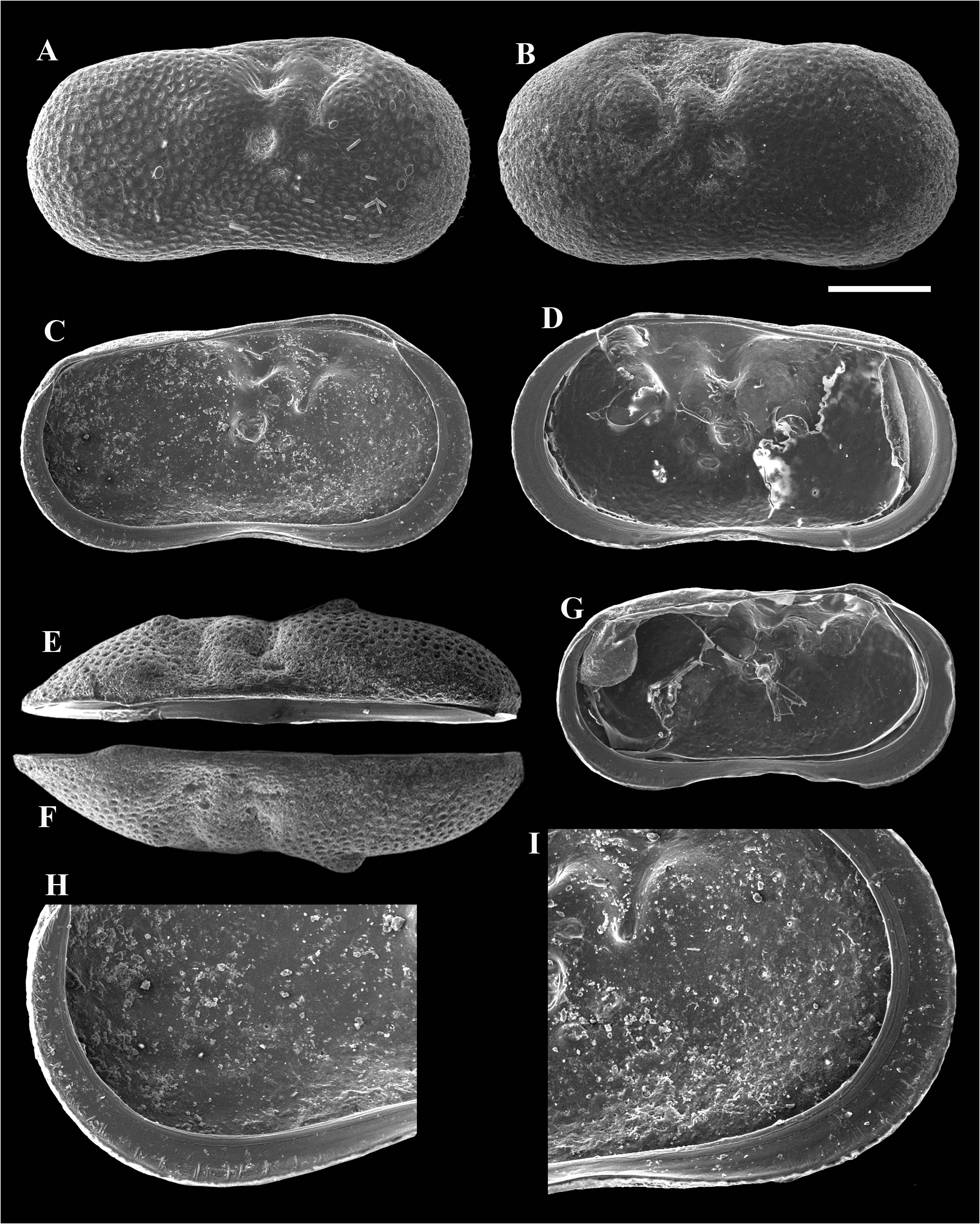
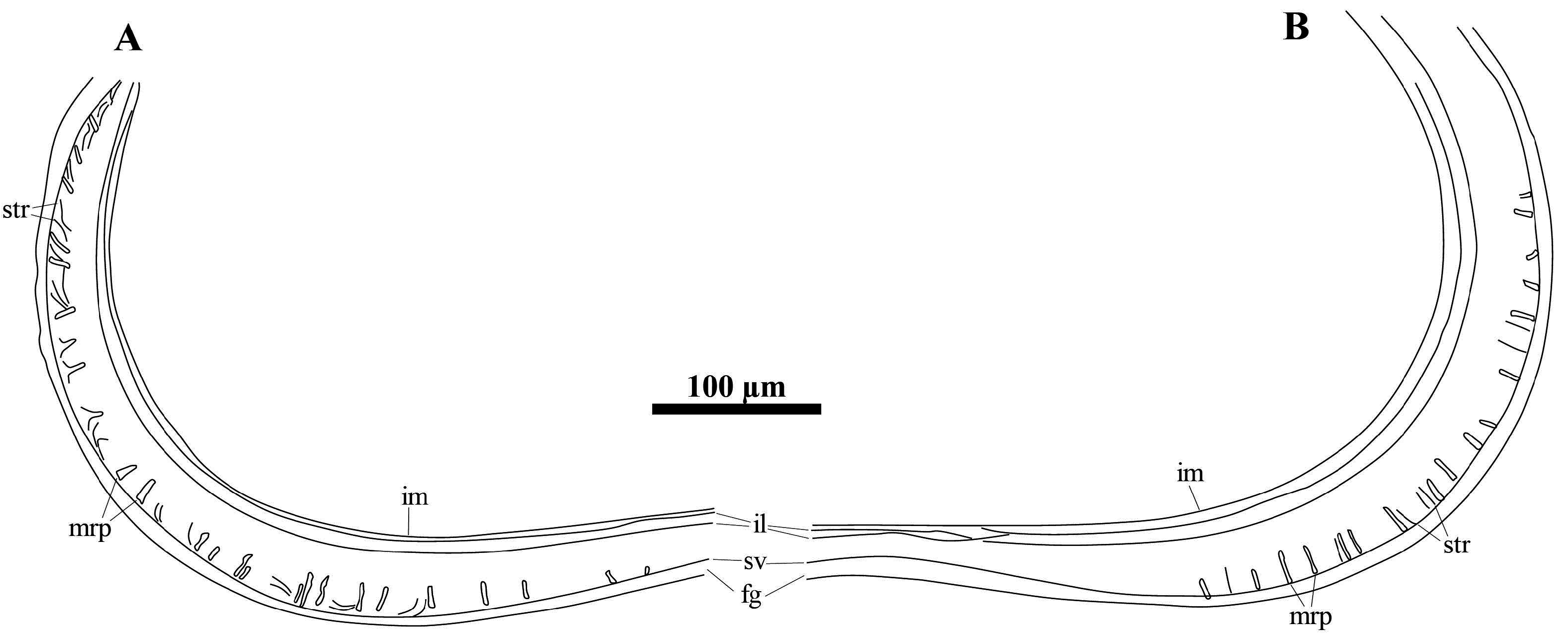
Description. Carapace ( Figs 18 View FIGURE 18 & 19 View FIGURE 19 ) sub-rectangular in lateral view. Dorsal margin nearly straight, only slightly concave at immediately posterior of mid-length. Highest point situated at 1/4 of valve length from anterior end. Anterior margin broadly rounded. Posterior margin slightly less arched than anterior margin. Ventral margin concave. Anterior sulcus situated at ca. 1/3 of valve length from anterior. Posterior sulcus situated at mid-length. Carapace sub-elliptical in dorsal view. Anterior end bluntly pointed, posterior end more rounded. Greatest width situated immediately behind posterior sulcus. Shell surface densely covered with small, shallow, rounded pits and two tubercles, one in postero-ventral area, and one in anterior dorsal margin, immediately anterior of sulcus. LV with more than 25 short marginal ripplets and number of striations in distal area of calcified inner lamella of posteroventral margin, and more than 15 short, weakly developed marginal ripplets and number of striations along calcified inner lamella of antero-ventral margin.
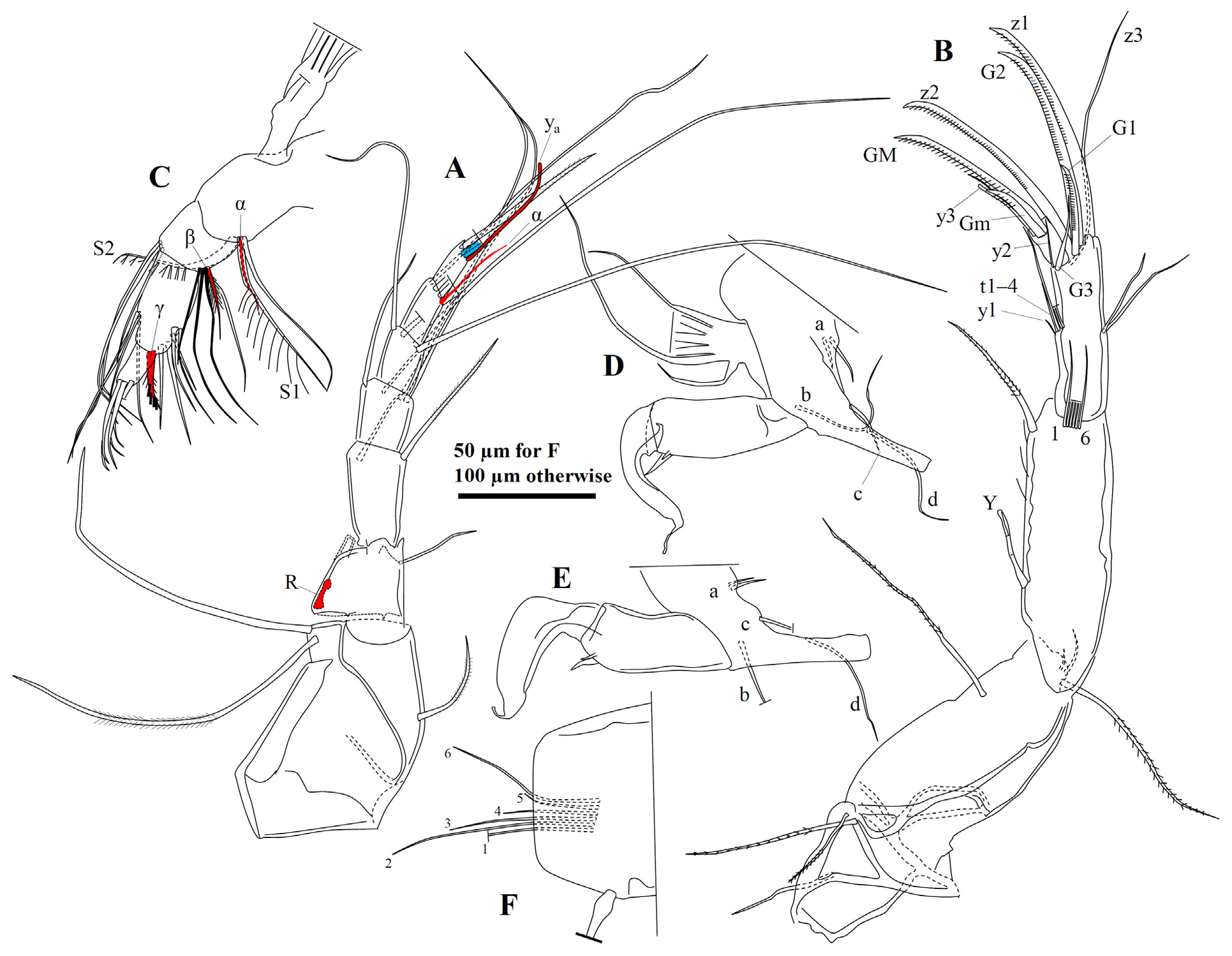
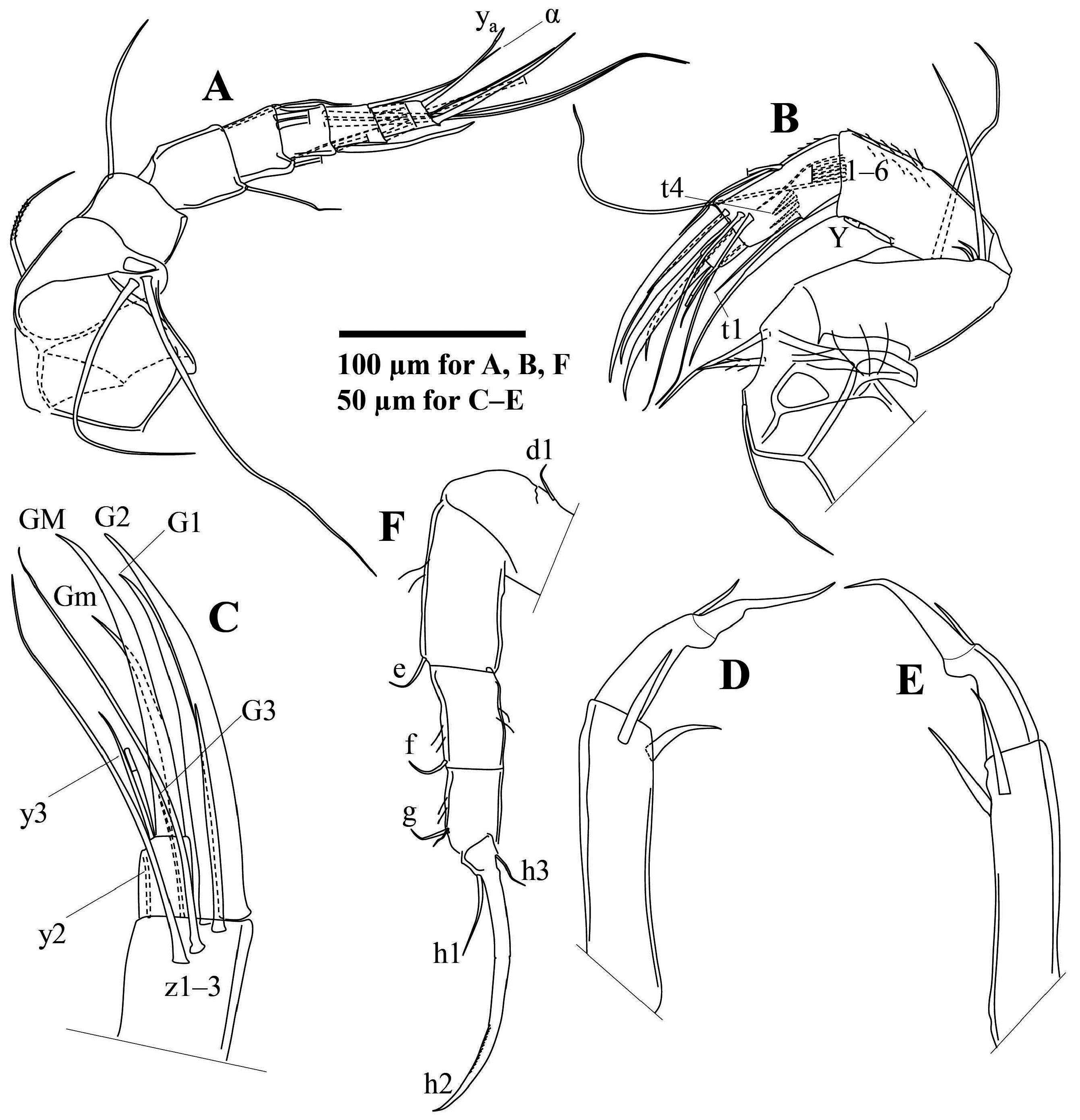
A1 ( Fig. 20A View FIGURE 20 ) with eight segments, first two of which fused, carrying one slightly setulous dorsal seta and two unequally long, smooth ventral setae. Third segment short and stout with one dorso-apical seta extending beyond fourth segment. Fourth segment sub-squarish with one fine dorso-apical seta extending to mid-length of seventh segment and one fine ventro-apical seta reaching mid-length of sixth segment. Fifth segment quadrate with two long dorso-apical setae extending beyond terminal segment with about half of length and two shorter, unequal ventroapical setae, one of which reaching beyond terminal segment, other one to terminal end of seventh segment. Sixth segment quadrate with two dorso-apical setae extending well beyond terminal segment and two ventro-apical setae, one of which extending beyond terminal segment with 1/5–1/4 of length, other one shorter and claw-like. Seventh segment sub-quadrate with five apical setae, shortest one (seta α) situated at exterior side of segment and extending almost to tip of aesthetasc y a. Eighth segment slightly elongate with three apical setae and aesthetasc y a situated at dorsal corner, shorter than claw-like seta at ventral corner. Two medial setae unequal in length.
A2 ( Fig. 20B, C View FIGURE 20 ) penultimate segment undivided in both sexes. First segment with one ventro-basal seta and two ventro-apical setae, shorter one of which sparsely plumose. Second segment with one ventro-apical seta extending to or slightly beyond mid-length of penultimate segment. Longest exopodal seta reaching distal end of third segment (first endopodal segment). Aesthetasc Y situated at mid-length of third segment, nearly reaching terminal end of third segment (first endopodal segment). Five long swimming setae on third segment reaching terminal claws or only slightly beyond, while sixth one only beyond middle of penultimate segment. Seta t1 extending somewhat beyond terminal segment. Setae t2 to t4 shorter, sub-equal in length, not reaching to end of segment. Seta z1 of male short, slightly claw-like. Setae z2 and z3 not transformed, extending to tips of claws. Female z-setae untransformed. G2 longest and most robust claw. G1 slightly shorter than G2. Male G3 very thin. Female G3 claw-like. Aesthetasc y2 very fine, reaching end of terminal segment. Male terminal segment with GM claw reaching tip of G2. Gm slightly shorter than GM. Aesthetasc part of y3 ca. 1/3 length of GM.
Md and Mx (not shown) typical of genus, with few specific features. Vibratory plate of Mx with four robust reflexed setae.
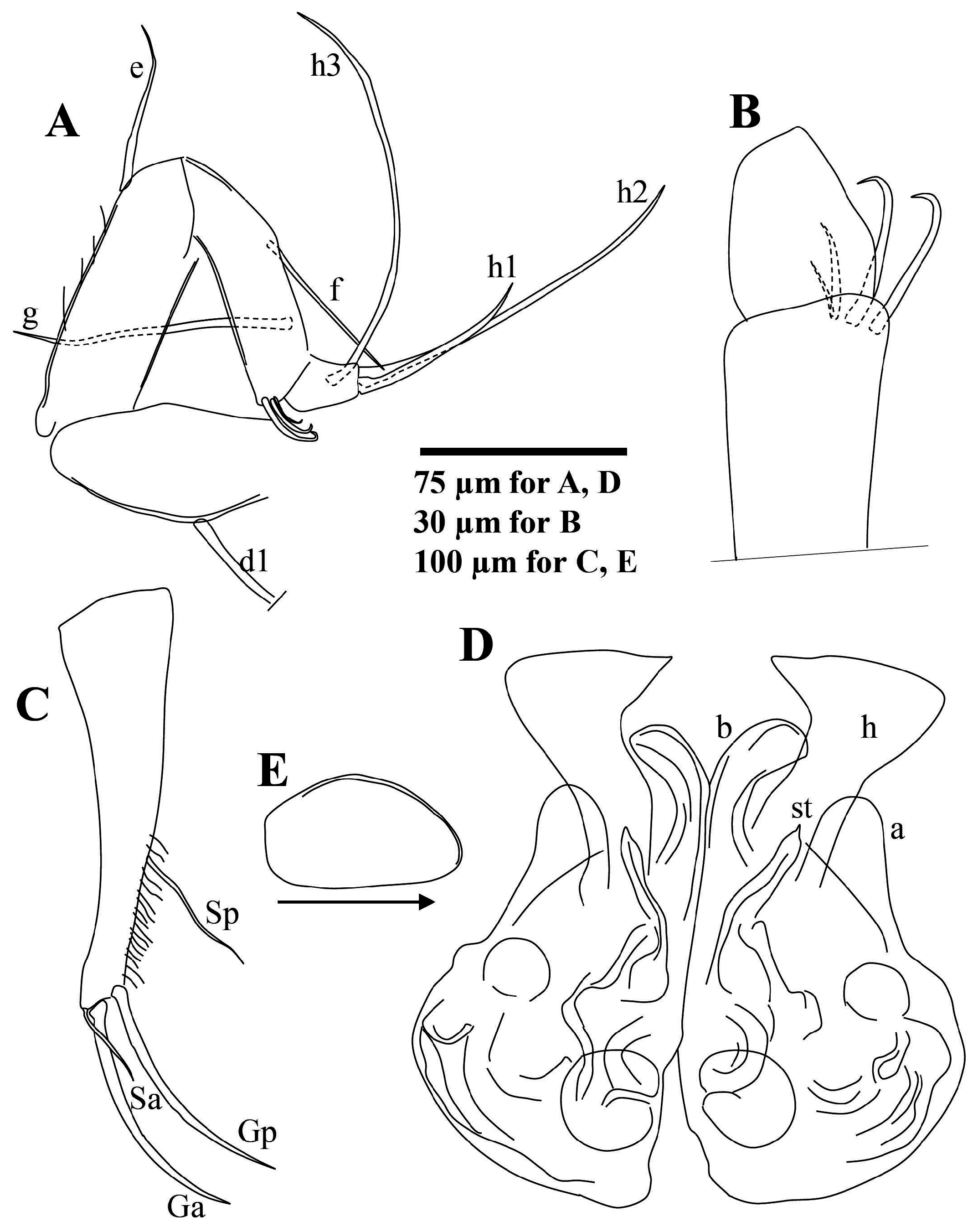
Male L5 palps ( Fig. 20D, E View FIGURE 20 ) sub-equal, 2-segmented. Basal segment (i.e., trunk) elongate with two ventro-apical setae. Terminal segment (i.e., finger) narrower at middle, with one robust apical seta and one short dorso-subapical seta.
L6 ( Fig. 20F View FIGURE 20 ) five-segmented. Seta d1 short, d2 absent, e, f and g sub-equal in length. Seta h1 longer than h3. Claw h2 faintly serrated along distal part.
L7 ( Fig. 21A, B View FIGURE 21 ) with four segments. Seta d1 robust, somewhat claw-like. Setae e and f sub-equally long, shorter than g. Penultimate segment apically with group of small, hook-like structures, two large, one intermediate, and one small sized. Seta h1 shorter but more robust than h2. Seta h2 shorter than h3.
Ur ( Fig. 21C View FIGURE 21 ) ramus comparatively robust for genus. Sp situated at ca. 2/3 length of ramus, extending to base of claws. Distal half of ramus with fine, long pseudochaetae. Ga and Gp slightly longer than half of ramus length.
Hp ( Figs 21D View FIGURE 21 & 22A View FIGURE 22 ) inner lobe (b) elongate and slender, distally curved towards middle lobe. Middle with relatively narrow neck distally widening to sub-triangular distal part, distal inner corner sharply angled, outer edge angled to rounded. Outer lobe (a) short, distally evenly rounded and partly overlapping middle lobe. Copulatory process (st) slender, with pointed end.
ZO ( Fig. 22D View FIGURE 22 ) with ca. 16–17 internal rosettes of spines, both ends inflated but proximal end much larger.
Female reproductive organ ( Fig. 21E View FIGURE 21 ) with one elongate, ovate lobe.
Remarks. The five-segmented L6, inflated proximal end of the ZO, and ‘blade-type’ copulatory process of the Hp indicates that Ilyocypris tibeta n. sp. belongs to the Ilyocypris japonica Okubo, 1990 group of Asian species as defined by Smith et al. (2019). The new species is particularly similar to Ilyocypris japonica , Ilyocypris mongolica Martens, 1991 and Ilyocypris innermongolica Zhai & Xiao, 2013 in terms of carapace, L7 (with apical hook-like structures on the third segment) and Hp morphologies ( Okubo 1990; Martens 1991; Zhai & Xiao 2013; Smith et al. 2019). Differences include the following: The A2 natatory setae of Ilyocypris tibeta n. sp. reach to about the tips of the terminal claws, while these are much longer in the other three species extending significantly beyond the terminal claws by almost 1/2 of their lengths. The setae on the three distal-most segments of the A1 of Ilyocypris tibeta n. sp. are also much shorter than those of I. japonica , I. mongolica and I. innermongolica ( Smith et al. 2019; D. Zhai, unpublished data; Zhai & Xiao 2013). The copulatory process (st) of the Hp of Ilyocypris tibeta n. sp. is distally more slender than those of both I. mongolica and I. japonica , and most similar to I. innermongolica . The outer lobe (a) is small, narrow and evenly rounded in Ilyocypris tibeta n. sp., while more angular in I. innermongolica and wider in both I. japonica and I. mongolica . The Hp middle lobe (h) of Ilyocypris tibeta n. sp. is similar to those of I. japonica and I. mongolica , although this lobe is slightly more rounded on the outer distal edge and less inflated in Ilyocypris tibeta n. sp. The Hp middle lobe of I. innermongolica is significantly different to Ilyocypris tibeta n. sp. and the other two species, being distally much less inflated. The ZO of all the male I. mongolica specimens (Zhai & Zhao 2014) have 13–14 rows of spines; while the number is larger in Ilyocypris tibeta n. sp. (16–17), similar to I. japonica (15–17) and I. innermongolica (17–21). The hook-like structures of the L7 are well-developed in Ilyocypris tibeta n. sp., similar to those of I. mongolica , and more pronounced than those of I. japonica and I. innermongolica .
The type locality of Ilyocypris tibeta n. sp. is a small, shallow pond with a soft, yellowish substrate, a few macrophytes and animal footprints around the edges. It was also found in two other places, a pond with a soft mud substrate, and in the riffle zone of a slowly flowing river ( Table 1 View TABLE 1 ). Altitudes of these sites ranges from 4325 to 4593 m.
Several reports of Ilyocypris species from the Tibetan Plateau figure valves that are very similar to Ilyocypris tibeta n. sp. (e.g. Wrozyna et al. 2009a; 2009b; Zhang et al. 2013; Song et al. 2015a; Akita et al. 2016; see synonymy list above). The carapaces of Ilyocypris species generally show a strong resemblance to each other, but can also show high amounts of intraspecific variation (e.g. tubercle development, and lateral view of the carapace), complicating identification on valves alone (e.g. Mazzini et al. 2014; Smith et al. 2019). The most reliable characters for identification are found on the appendages and male sexual organs. Thus it is not possible to determine if valves previously figured are definitely conspecific with Ilyocypris tibeta n. sp., and highlights the importance of soft part examination when possible.
The scattered known localities of living (and possible conspecific sub-fossil) material of this species suggests that it could be widespread at least in the southern and western parts of Tibet, and it probably favours shallow, muddy habitats.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Cypridoidea |
|
Family |
|
|
SubFamily |
Ilyocypridinae |
|
Genus |