
Ilyocypris incus, Smith & Zhai & Chang, 2019
|
publication ID |
https://doi.org/10.11646/zootaxa.4652.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:B8EA4D0C-07CC-4C8F-9B21-C97B9852AAFF |
|
DOI |
https://doi.org/10.5281/zenodo.5610194 |
|
persistent identifier |
https://treatment.plazi.org/id/A90187A1-BA52-FF88-128B-D51748D4F96B |
|
treatment provided by |
Plazi |
|
scientific name |
Ilyocypris incus |
| status |
sp. nov. |
Ilyocypris incus sp. nov.
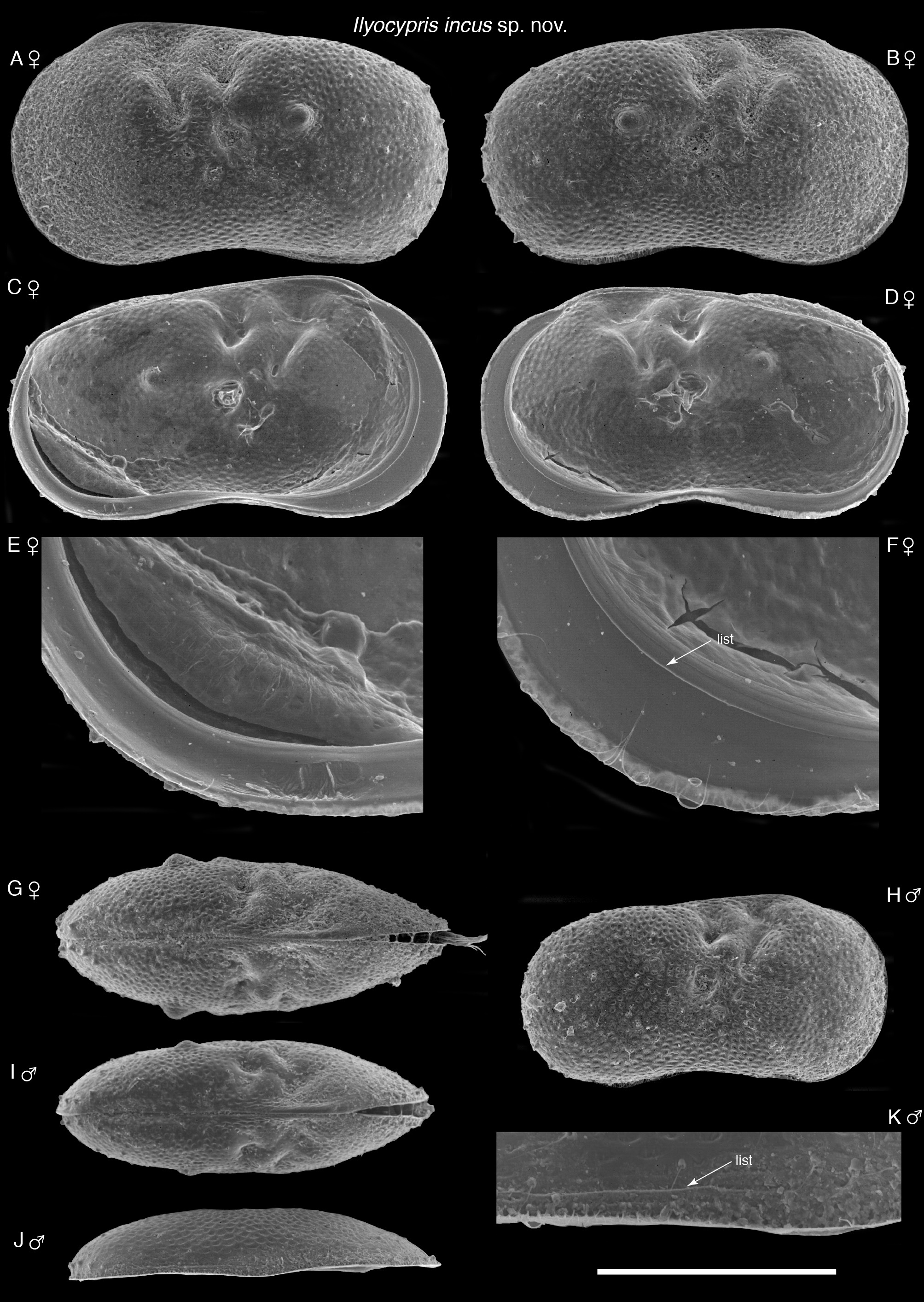
( Figs 5F View FIGURE 5 , 6C View FIGURE 6 , 14–17 View FIGURE 14 View FIGURE 15 View FIGURE 16 View FIGURE 17 )
1974 Ilyocypris angulata Sars, 1903 —Okubo: 46–50, fig. 1a–v.
? 1991a Ilyocypris dentifera Sars, 1903 —Kim & Min: 317–321, figs 8 & 9.
2004 Ilyocypris dentifera Sars, 1903 —Okubo: 16–17, figs 4m–q.
? 2012 Ilyocypris dentifera Sars, 1903 —Chang et al.: Table 1 View TABLE 1 (referring to Kim & Min 1991a).
2013 Ilyocypris angulata Sars, 1903 —Karanovic & Lee: 41, 55, 59, 64, fig. 14 (in part, referring to Okubo 1975 [sic]).
? 2014 Ilyocypris dentifera Sars, 1903 —Smith et al.: 1 (referring to Kim & Min 1991a).
Etymology. From the Latin ‘incus’ meaning anvil and referring to the shape of the copulatory process of the hemipenis (treated as a noun in apposition).
Diagnosis. Carapace quadrate in lateral view, covered in small pits, and approximately seven tiny spines near posterior margin. Anterior inner calcified lamellae of both valves with lists. With or without tubercles on carapace. Swimming setae of antenna very long, extending significantly beyond end of claws. Sixth limb with four segments (second endopodal segment undivided). Seventh limb with two setae on third segment (= second endopodal segment). Anterior end of Zenker organ small, slightly wider than central tube. Inner lobe of hemipenis longer than outer lobe, touching or slightly overlapping middle lobe, with bulbous end. Outer lobe nearly touching to slightly overlapping middle lobe, copulatory process well developed, with large, roughly triangular distal end with uneven distal edge.
Type locality. Rice fields in Bomun-dong, Gyeongju-si, Gyeongsangbuk-do, South Korea, 35.8364º N, 129.2483º E, altitude 73m.
Type material. Holotype: Male ( LBM 1430009132 View Materials ), dissected with appendages sealed in a glass slide and valves stored dry in a micropalaeontological cavity slide.
Allotype: Female ( LBM 1430009134 View Materials ), dissected with appendages sealed in a glass slide and valves stored dry in a micropalaeontological cavity slide.
Paratypes: Males ( LBM 1430009131 View Materials and DB40054), dissected with appendages sealed in glass slides and valves stored dry in micropalaeontological cavity slides. Male ( LBM 1430009138 View Materials ) whole carapace, stored dry in a micropalaeontological cavity slide .
Females ( LBM 1430009136 and DB40055), dissected with appendages sealed in glass slides and valves stored dry in micropalaeontological cavity slides. Female ( LBM 1430009137), whole carapace, stored dry in a micropalaeontological cavity slide.
All type material collected from the type locality on 4 July 2011.
Other material. Okubo (1974) stated that some specimens of the Japanese “ I. angulata ” he studied would be deposited in the collections of the National Science Museum, Tokyo, but enquiries revealed that the specimens are not in their collections.
Description. Male carapace length 746–797 µm, height 404–436 µm, height/length 0.53–0.55. Female carapace length 827–910 µm, height 463–504 µm, height/length 0.55–0.57. Anterior margin more inflated than posterior margin ( Fig. 14 View FIGURE 14 ). Dorsal margin almost straight, sloping down posteriorly, maximum height anterior of muscle scars. Ventral margin strongly concave. Well-defined bifurcated sulcus near dorsal margin just anterior of adductor muscle scars, which form small rounded depression below posterior part of sulcus. Surface of valves densely covered with many small pits. Area near posterior margin with about seven tiny spines. Both valves with faint outer lists running near ventral margins ( Fig. 14J & K View FIGURE 14 ). Tubercles present in some specimens, one each side of sulcus (posterior one more developed), one slight one in mid position of sulcus, one in postero-ventral region and one in central ventral region. Inner calcified lamella narrow, especially posteriorly. Anterior inner calcified lamellae of both valves with lists. Postero-ventral region of calcified inner lamella of left valve with shallow depression containing 2–3 marginal ripplets ( Fig. 14E View FIGURE 14 ). Adductor muscle scars obscured. Dorsal view oval, with posterior margin more rounded than anterior margin.
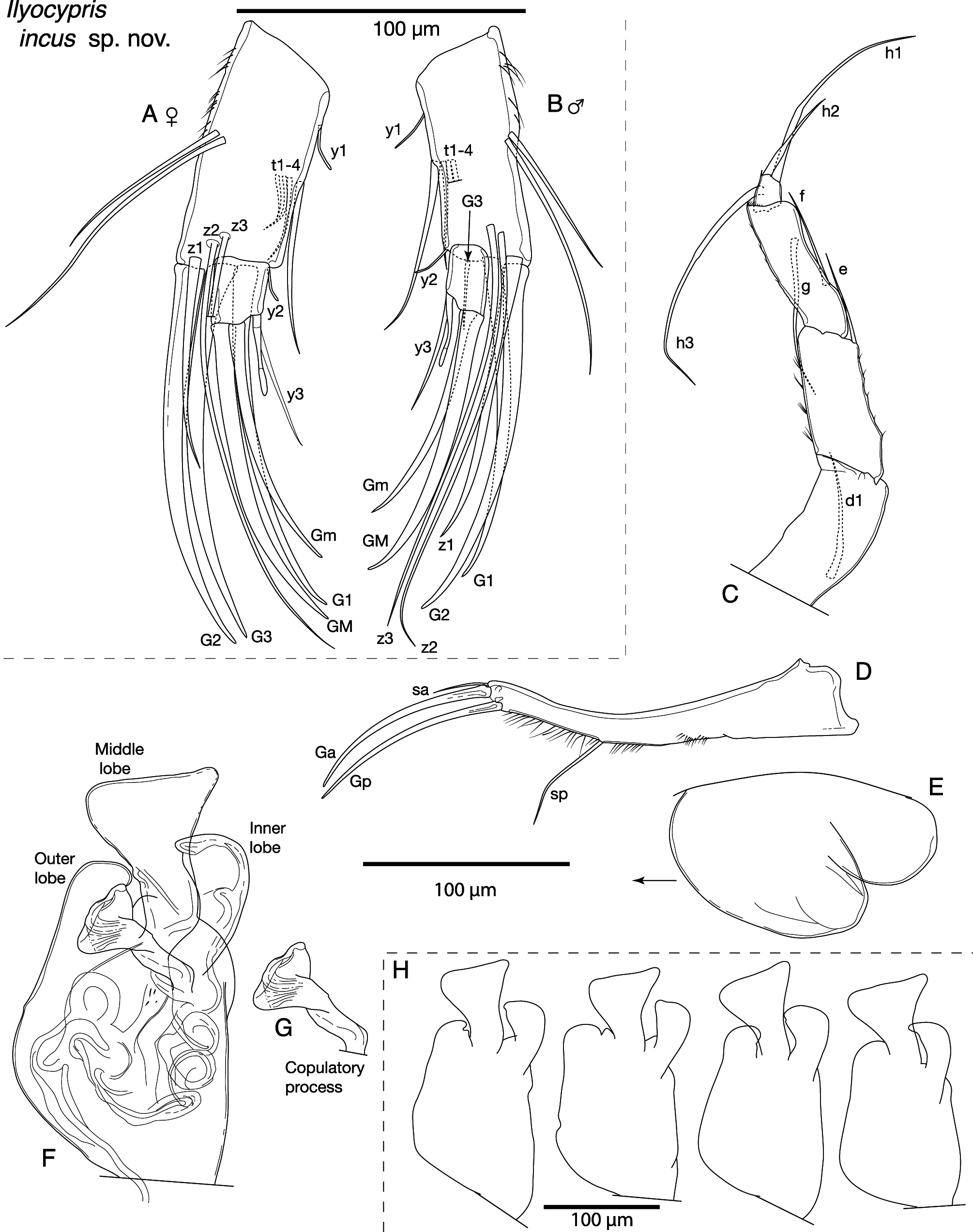
Antennule with seven articulated segments ( Fig. 15A View FIGURE 15 ). First segment with two long sub-apical setae on ventral margin and one medium-length, stout seta on dorsal margin. Second segment with relatively long apical-dorsal seta. Third segment with one short apical-dorsal seta and one short apical-ventral seta. Fourth segment with two long apical-dorsal setae, and two shorter apical-ventral setae. Fifth segment with two long apical-dorsal setae, and two shorter apical-ventral setae. Sixth segment with four long setae and relatively long alpha seta. Terminal segment apically with two short setae, one long seta and relatively short aesthetasc ya.
Male antennae with fourth segment undivided ( Fig. 15B View FIGURE 15 & 16B View FIGURE 16 ). Swimming setae very long, extending considerably beyond end of claws; shorter accompanying seta also long, reaching to mid-point of claws. G3 reduced to short seta, extending to distal end of terminal segment. Claw z1 small and slender, not reaching to end of claw G2. Female antenna with G2 claw as long as claws G1 and G3 ( Fig. 16A View FIGURE 16 ). Seta z1 stout, claw-like, shorter than setae z2 and z3, approximately half length of G2.
Mandibular coxa small, with well-developed teeth on endite ( Fig. 15D View FIGURE 15 ). Mandibular palp four-segmented ( Fig. 15 C View FIGURE 15 ). Alpha seta on first segment small and slender. Second segment with two setae on outer edge, and 3+1+beta setae on inner edge. Beta seta short, stubby and hirsute. Third segment with four setae on outer apical corner, three setae on inner apical corner, and one apical seta between other two groups. Terminal segment with three stout apical setae, two thinner and shorter apical seta and one seta protruding from area proximal of mid-length, towards outer edge.
Maxillula palp first segment with three sub-equal medium-length setae on outer apical corner, one mediumlength seta positioned on apical edge offset from apical corner, and one sub-apical setae in mid-width position ( Fig. 15E View FIGURE 15 ). Second segment wider than long, apically with three claw-like setae and three shorter setae.
Female fifth limb with large basis supporting two a-setae and approximately 17 setae of various lengths on inner edge (endite) ( Fig. 15F View FIGURE 15 ). Palp with two segments, terminating with one long and two mid-length setae. Branchial plate obscured, but with at least five rays. Male palp with long, thin proximal segment, supporting distally two setae and large grasping hook. Left and right hooks similar in morphology, gently curved and tapering to sharp tips, with short, distally very fine seta on outer edge ( Fig. 15H View FIGURE 15 ).
Sixth limb four-segmented ( Fig. 15G View FIGURE 15 ). First segment (= protopodite) with short d1 seta. Second segment (= first endopodal segment) with short e seta. Third segment (= second endopodal segment) with short f seta protruding from mid-length and short apical g seta. Terminal segment (= third endopodal segment) with relatively long h1 seta, short h3 seta and well-developed h2 claw, approximately 1.9x length of anterior sclerotized margin of first endopodal segment (marked with black triangle on Fig. 15G View FIGURE 15 ).
Seventh limb with four segments ( Fig. 16C View FIGURE 16 ). First segment with seta d1. Second segment with seta e reaching to approximately mid-length of third segment. Fourth segment with two setae on anterior edge (f and g), distal-most one longest. Fourth segment small, with seta h3 slightly longer than h1, and seta h2 short.
Caudal ramus typical of genus, ramus distinctly curved distal of mid-length ( Fig. 16D View FIGURE 16 ). Claw Gp slightly longer than Ga, seta sp long.
Female reproductive organ with large, rounded genital lobe anteriorly, and smaller rounded lobe posteriorly, just anterior of base of caudal ramus ( Fig. 16E View FIGURE 16 ). Organ delicate and easily distorted or damaged during dissections.
Hemipenes, outer lobe unevenly rounded distally, nearly touching to slightly overlapping middle lobe ( Fig. 5F View FIGURE 5 , 16F & H View FIGURE 16 ). Middle lobe triangular, inner corner sharply angled, outer edge more rounded. Inner lobe longer than outer lobe, with inner edge slightly curving along most of length, before smoothly curving to bulbous distal end; bulbous end projecting towards middle lobe, sometimes slightly overlapping. Copulatory process well developed, with large, roughly triangular distal end with uneven distal edge ( Fig. 16G View FIGURE 16 ).
Zenker organ with 18–19 internal rosettes ( Fig. 6C View FIGURE 6 ). Proximal end oval and small, slightly wider than central tube.
Intraspecific variation. Some specimens have tubercles developed on the carapace to varying degrees, while others have no obvious tubercles. Okubo (1974; as I. angulata ) figured specimens with five tubercles on each valve. Seta sa on the caudal ramus is variable in length, with that of some specimens up to 157% longer compared with the holotype’s. The inner lobe of the hemipenis is rather variable in appearance distally, sometimes with a slight gap between it and the middle lobe, while in other specimens slightly overlapping the middle lobe ( Fig. 16H View FIGURE 16 ).
Remarks. This species is very similar to I. dentifera , I. hanguk , I. dui , and I. gibba . The valves are more round- ed posteriorly than those of I. dentifera , which tends to have a more truncated posterior. The hemipenes demonstrate several differences, including the shapes of the outer and inner lobes, and the copulatory process. Our specimens are a close match to the incorrect records of I. angulata and I. dentifera figured by Okubo (1974; 2004), and we consider them conspecific.
The misidentified Ilyocypris dentifera of Kim & Min (1991a) comes close to this species, but the outer lobe of the hemipenis has a slightly different shape and the claw of their figured sixth limb is robustly serrated. The outer lobe of the hemipenis does vary somewhat in our material, and so the hemipenis morphology recorded by Kim & Min (op. cit.) could represent the same species. The robust serration on the sixth limb is unusual for Ilyocypris species generally, and maybe a result of Kim & Min’s drawing style; other species figured by them also have robust, rather exaggerated serration on the sixth limbs ( Kim & Min 1991a; 1991b). We consider the records of I. dentifera of Kim & Min (1991a) at least in part, as possibly conspecific with Ilyocypris incus sp. nov.
Ilyocypris hanguk is similar to Ilyocypris incus sp. nov., but the posterior margin of the male carapace is more rounded in the latter (adult female morphology of I. hanguk remains unknown). The three lobes on the hemipenis are different in shape compared with those of Ilyocypris incus sp. nov., in particular, the middle lobe of I. hanguk is proximally wider and the copulatory process is missing the large distal projections found in Ilyocypris incus sp. nov. Other features of I. hanguk that are different include the very small h1 seta on the walking leg (note that the h1 and h3 setae of the sixth limbs are mislabelled in Karanovic & Lee 2013), and the large number of rosettes in the Zenker organ (approximately 25 in I. hanguk , 18–19 in Ilyocypris incus sp. nov.).
Similar to I. hanguk , the carapace of I. dui resembles that of I. dentifera more than Ilyocypris incus sp. nov. The middle lobe of the hemipenis of I. dui is most similar to that of I. hanguk in that it is proximally wide, which also distinguishes it from Ilyocypris incus sp. nov. The outer and inner lobes and the copulatory process are also differently shaped than those of Ilyocypris incus sp. nov.
Ilyocypris gibba has a similar outline in lateral view to Ilyocypris incus sp. nov. and overall the differences in carapace morphology are subtle. Ilyocypris gibba has more ripplets (6–10) on the postero-ventral calcified inner lamella of the left valve than Ilyocypris incus sp. nov. (2–3), but the numbers of ripplets are known to vary within species and are not considered to be a reliable taxonomic feature ( Mazzini et al. 2014). The pitting on the carapace is coarser and the postero-dorsal corner more angular in I. gibba compared with Ilyocypris incus sp. nov. The appendages are also very similar in the two species, with only the hemipenes and Zenker organs showing clear differences; the inner lobe of the hemipenis of I. gibba is much shorter and the outer lobe more angular than those of Ilyocypris incus sp. nov. ( Meisch 2000). The shape of the copulatory process of I. gibba remains unknown. The Zenker organ of I. gibba has a more inflated proximal end compared with that of Ilyocypris incus sp. nov.
Several species known from China don’t have appendages described, so comparisons with these species rely solely on the carapace morphology. Ilyocypris xizangensis Yang, 1982 (in Huang et al. 1982) is noticeably more elongate, and Ilyocypris sebeiensis Yang et al., 2004 a lot less elongate compared to Ilyocypris incus sp. nov. Ilyocypris echinata Huang, 1979 has lateral spines on the carapace, not seen in Ilyocypris incus sp. nov. Ilyocypris microspinata Huang, 1982 has a more sloping dorsal margin and a less inflated posterior in lateral view. The Pleistocene Ilyocypris opima Huang, 1982 is much more rounded in lateral view and Ilyocypris crassireticulata Huang, 1982 has much coarser pitting.
Distribution. Currently known from Gyeongsangbuk-do, South Korea, and Okayama Prefecture, Japan ( Fig. 9 View FIGURE 9 , see Table 1 View TABLE 1 for more details). Both known occurrences of this species fall into the temperate, without dry season, hot summer Köppen climatic zone (Cfa) ( Peel et al. 2007). Kim & Min (1991a) reported “ I. dentifera ”, which possibly represents this species, from various places throughout South Korea. However, considering the close similarity of this species with the true I. dentifera , it is unknown how many of these records potentially refer to Ilyocypris incus sp. nov.
Ecology. Ilyocypris incus sp. nov. was found in a rice field with I. dentifera and I. japonica during early July in Korea. Okubo (1974, as I. angulata ) reported that it is found from spring to summer, sometimes autumn, “creeping” in mud, although the habitat was not mentioned.





| LBM |
Lake Biwa Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Cypridocopina |
|
Family |
|
|
SubFamily |
Ilyocypridinae |
|
Genus |
Ilyocypris incus
| Smith, Robin James, Zhai, Dayou & Chang, Cheon Young 2019 |
Ilyocypris incus
| Smith & Zhai & Chang 2019 |
Ilyocypris incus
| Smith & Zhai & Chang 2019 |
Ilyocypris incus
| Smith & Zhai & Chang 2019 |
Ilyocypris incus
| Smith & Zhai & Chang 2019 |
Ilyocypris incus
| Smith & Zhai & Chang 2019 |
Ilyocypris incus
| Smith & Zhai & Chang 2019 |
Ilyocypris incus
| Smith & Zhai & Chang 2019 |
Ilyocypris sebeiensis Yang et al ., 2004 a
| Yang et al. 2004 |
Ilyocypris xizangensis
| Yang 1982 |
Ilyocypris microspinata
| Huang 1982 |
Ilyocypris opima
| Huang 1982 |
Ilyocypris crassireticulata
| Huang 1982 |
Ilyocypris echinata
| Huang 1979 |
Ilyocypris angulata
| Sars 1903 |
Ilyocypris dentifera Sars, 1903
| G. O. Sars-Chen 1903 |
Ilyocypris dentifera Sars, 1903
| G. O. Sars-Chen 1903 |
Ilyocypris dentifera Sars, 1903
| G. O. Sars-Chen 1903 |
Ilyocypris angulata
| Sars 1903 |
Ilyocypris dentifera Sars, 1903
| G. O. Sars-Chen 1903 |