
Heteranthus verruculatus Klunzinger, 1877
|
publication ID |
https://doi.org/ 10.1163/18759866-BJA10015 |
|
DOI |
https://doi.org/10.5281/zenodo.8356645 |
|
persistent identifier |
https://treatment.plazi.org/id/48500508-B37B-FFFD-E99F-B3B2FBABF915 |
|
treatment provided by |
Felipe |
|
scientific name |
Heteranthus verruculatus Klunzinger, 1877 |
| status |
|
Heteranthus verruculatus Klunzinger, 1877 View in CoL View at ENA figs. 2–10 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10
Heteranthus verruculatus Klunzinger, 1877: 84 View in CoL (original description); Carlgren, 1900: 92; Stephenson, 1922: 290; Carlgren, 1943: 31; Carlgren, 1949: 75; Cutress, 1977: 138; den Hartog, 1997: 358; Fautin et al., 2008: 52; Fautin, 2016: 416
Actinothrix verruculata: Andres, 1883: 509
Heteranthus verriculatus View in CoL [no author]: Carlgren, 1943: 32
Heteranthus insignis Carlgren, 1943: 30 (original description); Carlgren, 1950: 440; Fautin, 2016: 275 NEW SYNONYMY
Heteranthus sp. : González-Muñoz et al., 2015: 3; Brugler et al., 2018: 10
Material examined (*- observed alive by the first author) ( table S2 View TABLE 2 ). Indian Ocean. – Oman, Bar Al Hikan Peninsula ( UFM 4220 x2); Western Australia, Perth, Rottnest Island : Radar Reef ( WAM Z33600 x1; WAM Z33604 x6), Cape Vlamingh ( WAM Z33601 x3).
Western Pacific Ocean. – Singapore: Cyrene Reef (ZRC.CNI.0496 x2; ZRC.CNI.0505 x8; ZRC. CNI.0506 x3; ZRC.CNI.0507 x17; ZRC.CNI.0583 x3; ZRC.CNI.1207 x8*; ZRC.CNI.1230 x>50*; ZRC.CNI.1367 x18*), Pulau Jong (ZRC.CNI.1368 x>35*), Pulau Tekukor (photograph only), RafflesLighthouse(photographonly); Indonesia (Pulau Ambon): Latuhalat (RCLA.Cni.004 x3*), Tial (RCLA.Cni.003 x4*, RCLA.Cni.008 x2*); Eastern Australia: Low Isle on Snapper Island, Great Barrier Reef (NHM 1954.6.28.23 x1), Moreton Bay (MTQ-G59967 x2; MTQ-G59968 x3; MTQ-G59391 x21; MTQ-G59392 x1), Mystic Sands, Townsville (MTQ-G59393 x5).
South Pacific Ocean. – French Polynesian Islands, Mangareva, Rikitea (MNHN-IK-2019-8 x3).
Nomenclatural considerations and type material examined. Both Fautin et al. (2008: 52) and Fautin (2016: 416) listed two syntypes of H. verruculatus : the first is a nearly complete polyp that is kept in Berlin(ZMB 1852),while the second is a small fragment of a syntype, kept in Stockholm (NRS 4861). Regarding the latter, it has a recent label indicating that the sample was collected by ‘ Stuhlmann’ between ‘1888– 1889,’ from ‘ East-Africa , Sansibar, Kokotoni, S of Insel Puopo. Rock.’ This recent label is almost certainly an error. Despite these two syntype lots being kept in separate museums, they are not of different individuals; the fragment present in Stockholm is from the same individual present in Berlin, therefore only one syntype specimen exists instead of two.
While Carlgren’s (1900) publication dealt with material collected by Stuhlmann from East Africa, it also presented diagnoses of Heteranthidae and Heteranthus . Carlgren (1900: 92) did not report any species of the genus collected from that locality and explicitly states that, “ … Heteranthus KLUNZ. mit nur einer Spezies, H.verruculatus . Diese Spezies ist nicht in der STUHLMANN’schen Sammlung enthalten,”. In other words, Carlgren did not find Heteranthus individuals from East Africa in Stuhlmann’s collection. We infer that Carlgren’s (1900) inclusion of the diagnoses served to facilitate discussion regarding East African Phymanthus specimens, which he described in detail in his publication.
Older labels within lot NRS 4861 provided further support that this fragment is part of the syntype kept in Berlin. Though faded, one label has the number “1852,” written on it, which corresponds to the catalogue number present on the old label of the Berlin specimen ( fig. 2A View FIGURE 2 ). Other older labels, likely written by Carlgren himself, indicate that the fragment was from “ Röda havet ” [= Red Sea], and “Klunz orig!”. In addition, the NRS fragment corresponds to the position of the missing slice of ZMB 1852, though slightly smaller in width, with the oral disc and actinopharynx absent; presumably Carlgren had prepared histological slides from the NRS material (see Carlgren, 1950). This confirms that the H. verruculatus fragment numbered NRS 4861 is part of the syntype ZMB 1852.
We hereby designate both ZMB 1852 and NRS 4861, previously considered as separate syntypes but now found to be of the same syntype specimen, as the lectotype for the name Heteranthus verruculatus Klunzinger, 1877 (Code Article 74), to enhance its nomenclatural objectivity. While C.B. Klunzinger described the species from more than one individual that he had collected from Egypt ( Klunzinger, 1877; Fautin, 2016), the first author was not able to locate any other syntypes at museums where Klunzinger’s specimens are now kept, or in those that O. Carlgren have visited and possibly transferred some. Should these other syntypes be found, they are the paralectotypes of H. verruculatus (Code Article 74.1.3). In this publication, all three conditions stipulating the designation of a lectotype after 1999 have been met (Code Article 74.7).
Heteranthus verruculatus – Lectotype. ZMB 1852 ( fig. 2A, B View FIGURE 2 ), collected by C.B. Klunzinger, no later than 1877 from the Red Sea ( Koseir ). Two pieces of a single specimen, rigid and brittle, cut transversely across at mid-column ( fig. 2B View FIGURE 2 ). Distalmost column slit longitudinally, a slice of distalmost margin missing ( fig. 2B View FIGURE 2 ). An oily scum layer encrusts oral disc. Both pieces dark-greenish or grey. Total length 14 mm, mid-column 13 mm wide, distalmost and proximal end 15 mm wide, respectively; NRS 4861 ( fig. 2C View FIGURE 2 ) a single fragment originating from ZMB 1852. A piece of distalmost end of column, also includes part of the mid-column, dark sandy-brown in colour, in good condition. Fragment 4 mm wide.
Heteranthus insignis – Holotype, NRS 4076 , collected by D.C. Dawydoff from Vietnam and given to O. Carlgren ( fig. 2D View FIGURE 2 ). A single specimen, 17 mm in length, cut transversely into two pieces at mid-column, with a longitudinal cut along each. A slice of distalmost margin missing. Both pieces firm, in good condition. Both pieces cream-coloured in preservative.
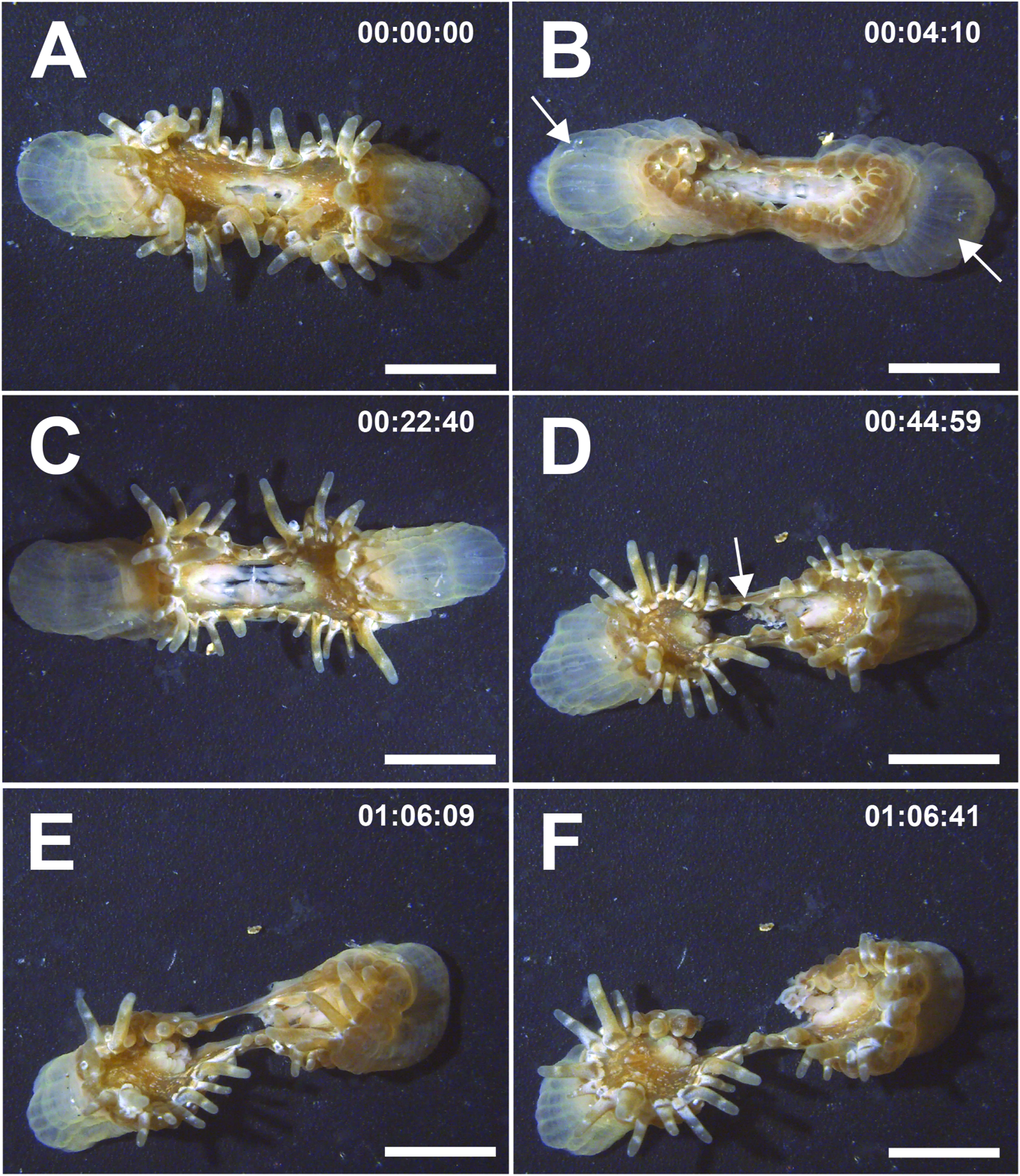
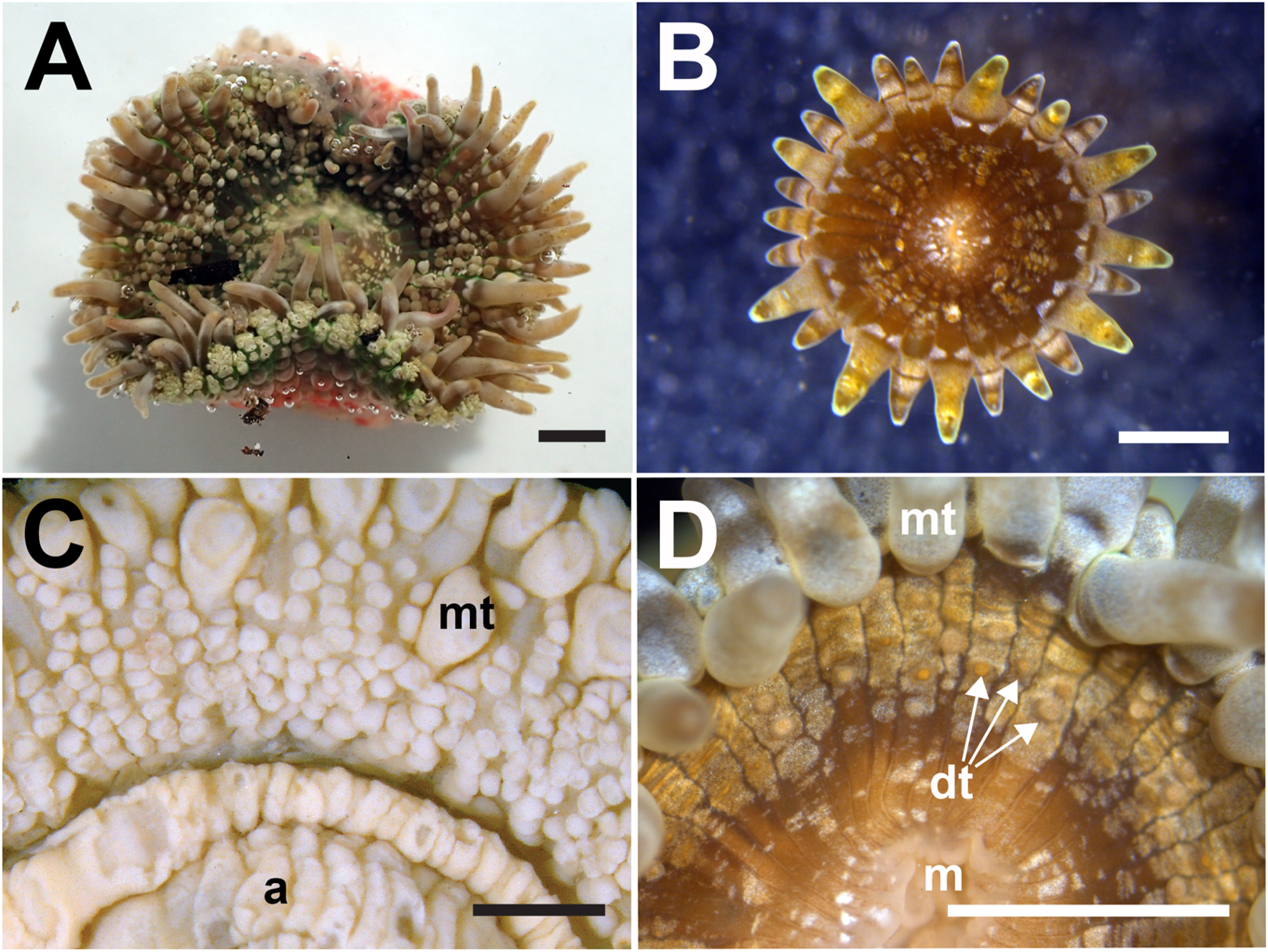
Natural history. Encountered during low spring tides along shallow tropical coasts, with oral disc and marginal tentacles expanded ( fig. 3A View FIGURE 3 ). Sand grains and shell fragments adhere to verrucae at distalmost end ( fig. 3B View FIGURE 3 ). May occur as solitary or as clonal, clustering individuals ( fig. 3C, D View FIGURE 3 ), inhabiting narrow rocky crevices. Clustering type abundant in sandy seagrass meadows of Halophila ovalis , with pedal end attached to the roots of plant or coral rubble underneath. Asexual propagation via longitudinal fission frequently observed ( fig. 4A–F View FIGURE 4 ;supplementary video S1).Zooxanthellate.
Oral disc. When fully expanded, margin may be slightly undulate in solitary individuals; in smaller clustering ones, flat( fig.5A, B View FIGURE 5 ).Outline oval to round; elongated oval during onset of longitudinal fission ( fig. 4A View FIGURE 4 ). In life, light to dark-brown, may be speckled with white patches. Thin-walled, mesenterial insertions seen as radial dark lines extending from mouth to marginal tentacles in living animals, as white lines in preserved materials. Discal tentacles present, papilliform ( fig. 5C, D View FIGURE 5 ). Area immediately around mouth without discal tentacles ( fig. 5D View FIGURE 5 ), arranged in radial rows from mid-way extending towards marginal tentacles. Radial rows of discal tentacle both endocoelic and exocoelic, numerous. In solitary individuals, discal tentacles conspicuous and densely arranged, clustering type less conspicuous and less dense ( fig. 5D View FIGURE 5 ), becoming obscure when preserved. Central mouth flat, edges tinged brown, white or pink in life, cream-white in preserved specimens.
Marginal tentacles. Numerous, numbers variable; in solitary individuals that exhibit a regular symmetry (i.e., mesenteries arranged symmetrically) typically 96, arranged in five cycles (e.g., RCLA.Cni.008). One per endo/exocoel; innermost cycles endocoelic, outermost exocoelic. Innermost cycle longer than outermost, longest length approximately ¾ radius of oral disc. Simple, conical, without ramified protuberances in life, some individuals slightly inflated at mid-tentacle ( figs. 3A, C View FIGURE 3 , 5A View FIGURE 5 ). Tip blunt, without perforation in live individuals; after preservation may appear perforated due to tentacle retraction (e.g., NHM 1954.6.28.23; fig. 5C View FIGURE 5 ). Wide at base, narrow toward tip. Colour in life greenishbrown, some with light horizontal cross bands along oral face ( fig. 5A, D View FIGURE 5 ). Base coloured white to dark brown, tip with white specks in life, cream-white in preserved specimens.
Column. Distalmost end flared slightly outwards when expanded. Distal margin with marginal projections, both endo/exocoelic. Larger marginal projections, endocoelic; smaller: exocoelic. In solitary individuals, marginal projections densely covered with conspicuous, papillae-like verrucae ( fig. 6A– C View FIGURE 6 ) that may be less dense or inconspicuous in clustering individuals (e.g., ZRC.CNI.1207, MTQG59393; fig. 6D View FIGURE 6 ). Longitudinal rows of round verrucae extend proximally from marginal projections, smaller near distalmost end, increasing in size proximally towards mid-column. Verrucae very conspicuous in solitary individuals, edges thickened and slightly raised, middle depressed ( fig. 6E View FIGURE 6 ); less conspicuous in clustering polyps ( fig. 6F View FIGURE 6 ). Longitudinal verrucae rows both endo/ exocoelic. Endocoelic rows longer, up to 20 verrucae per row; exocoelic: shorter, up to four in each row; shell fragments, coral rubble and/or sand grains may adhere to the verrucae. Column wall thin, mesenterial insertions can be seen through as faint light lines extending from distal to proximal end. From mid-column to proximal end of column smooth, without cinclides. Fosse present, shallow. In life, light-brown with greenishtinge to cream-white with reddish orange/ pink splotches, distalmost end dark grey ( fig. 6E–F View FIGURE 6 ). In preserved specimens, column entirely cream-white; distalmost end grey in recently preserved materials.
Pedal disc. Adheres readily to surfaces when animal is alive. Outline: oval, may be flat or inflated. Limbus slightly scalloped. Thin-walled, mesenterial insertions seen as light lines. Diameter may be equal or greater than oral disc.
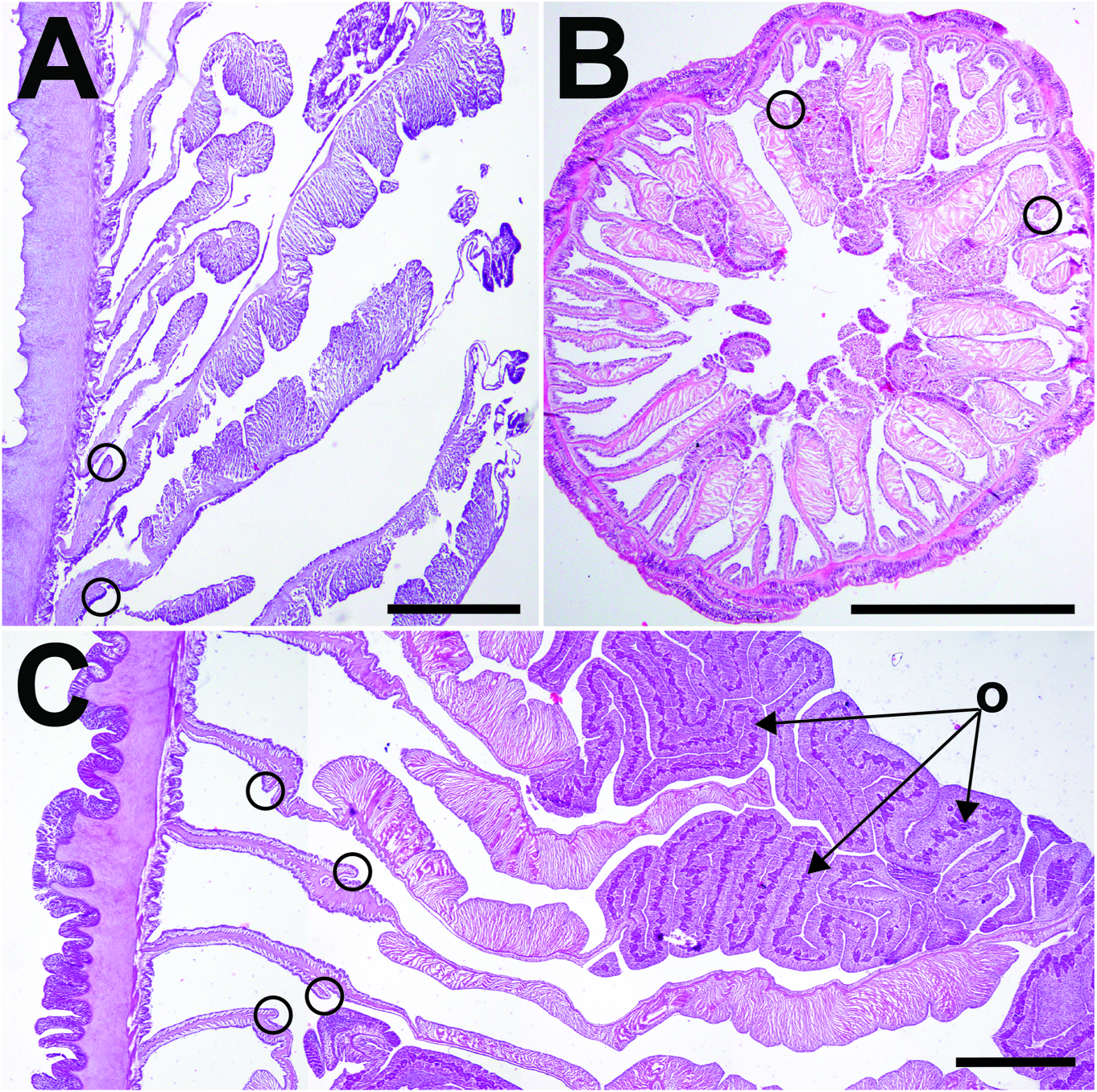
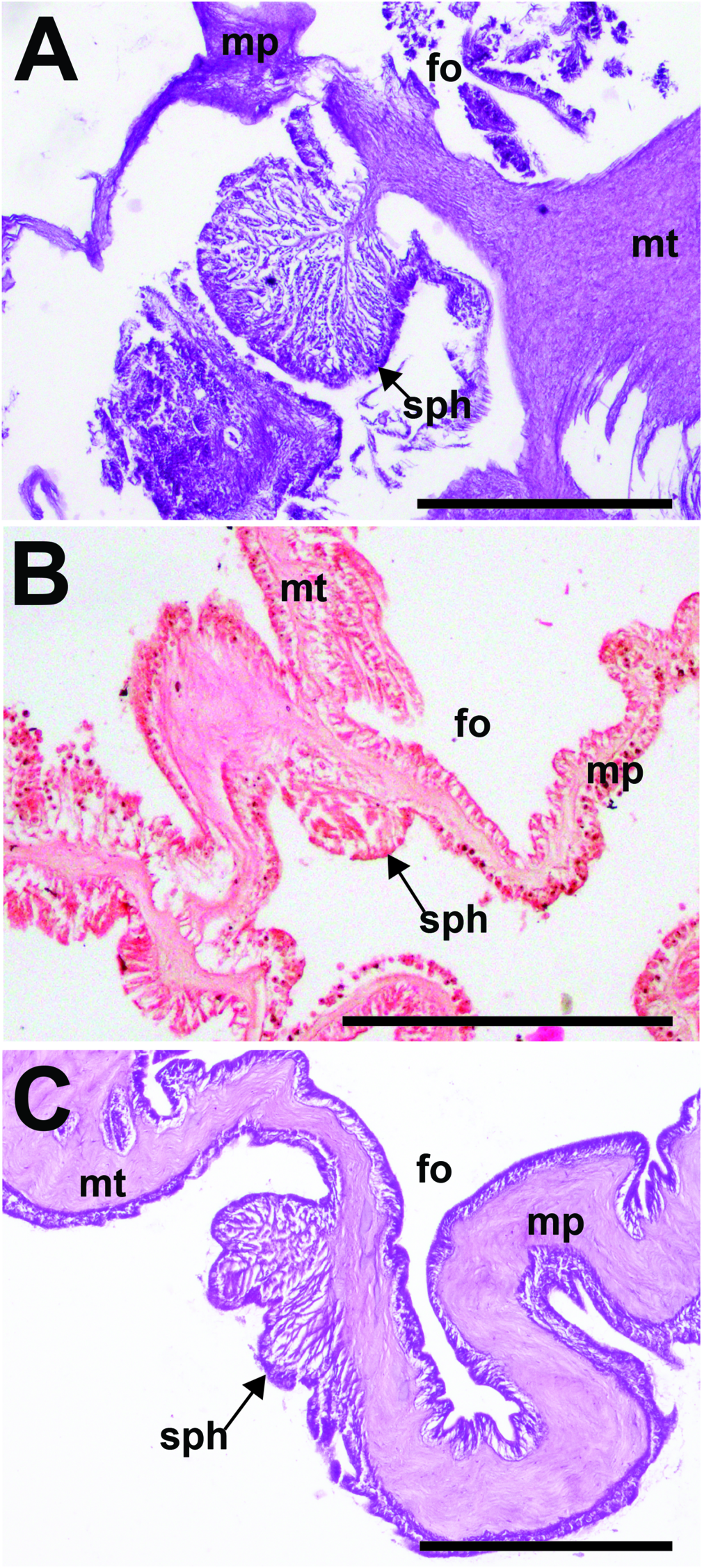
Internal anatomy. Anatomy typically arranged regularly in solitary individuals (e.g.,RCLA.Cni.008); irregular in clustering individuals (e.g., NRS 4076, WAM Z33601). Actinopharynx pleated longitudinally, extends proximally to mid-column; white in life, cream-colored in preserved specimens. Marginal and oral stomata present. Usually two siphonoglyphs in solitary individuals, symmetrical; clustering individuals with supernumerary siphonoglyphs (i.e., three siphonoglyphs in WAM Z33604) or may be entirely absent, asymmetrical. Mesenteries numerous, number of pairs variable in clustering individuals ( fig. 7A, B View FIGURE 7 ). In solitary individuals, mesenteries typically arranged in up to four cycles (i.e., 6 + 6 + 12 + 24 pairs), symmetrical. Mesentery pairs in first two cycles complete, those of third cycle incomplete, all with filaments and gametogenic tissue, if present ( fig. 7C View FIGURE 7 ). Mesentery pairs in fourth cycle incomplete, without retractor muscle or filaments. Retractor muscles strong, diffuse, always humped along its edge but less pronounced in clustering individuals. Parietobasiliar muscles poorly developed, retractor pennon reduced or absent; if present, extends away from mesentery ( fig. 7A–C View FIGURE 7 ). Directives attached to a siphonoglyph each, if present. Sphincter muscle below fosse, form variable among specimens. Typically strong, conspicuous and circumscribed, or restricted or with a few folds ( fig. 8A–C View FIGURE 8 ).
Cnidom Spirocysts, basitrichs, microbasic amastigophores, and microbasic p -mastigophores ( table 2 View TABLE 2 ; fig. 9 View FIGURE 9 ).
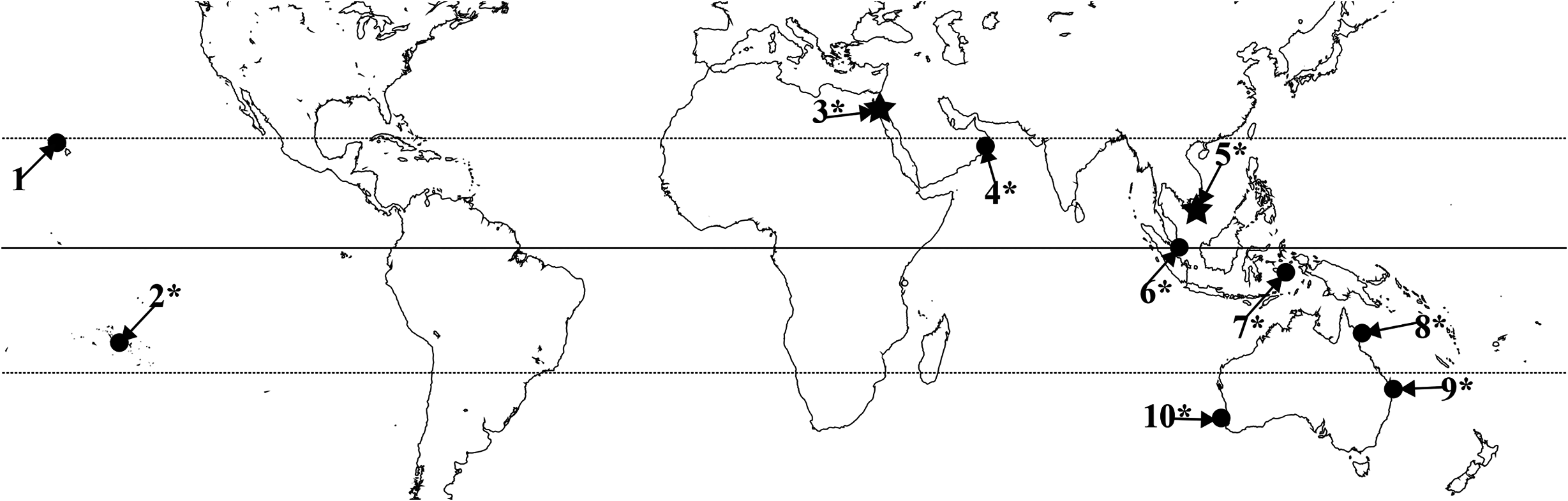
Distribution ( fig. 10 View FIGURE 10 ). Type localities: Egypt, Koseir (of H. verruculatus ; Klunzinger, 1877); Vietnam, Poulo Condore [= Côn SƠn Island, Vung Tau] (of H. insignis ; Carlgren, 1943). Published records: Eastern Australia, Great Barrier Reef ( Carlgren, 1950), Moreton Bay and Townsville ( Fautin et al., 2008); Indonesia ( den Hartog, 1997: locality not specified); Hawaii, Oahu, Kaneohne Bay ( Cutress, 1977); Western Australia ( Fautin et al., 2008: locality not specified). New records: French Polynesian Islands, Mangareva, Rikitea; Oman, Bar Al Hikman Peninsula; Singapore.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Heteranthus verruculatus Klunzinger, 1877
| Yap, Nicholas Wei Liang, Quek, Zheng Bin Randolph, Tan, Ria, Nugroho, Dharma Arif, Lee, Jen Nie, Lee Berumen, Michael, Tan, Koh Siang & Huang, Danwei 2021 |
Actinothrix verruculata:
| Andres 1883: 509 |
Heteranthus verruculatus
| Klunzinger 1877: 84 |
Heteranthus verriculatus
| Klunzinger 1877 |