
Grammia anna, (GROTE)
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2008.00496.x |
|
persistent identifier |
https://treatment.plazi.org/id/03B987FA-FFA7-F736-FF71-6DA9FE45E77F |
|
treatment provided by |
Felipe |
|
scientific name |
Grammia anna |
| status |
|
GRAMMIA ANNA (GROTE) (FIGS 9, 46, 102)
Arctia anna Grote, 1864a: 335 .
Arctia persephone Grote, 1864b: 433 .
Apantesis anna (Grote) ; Franclemont, 1983: 117. Grammia anna (Grote) ; Ferguson & Opler, 2006: 9.
Type material: Arctia anna : described from a female holotype, type no. 7685 in ANSP [not examined]; the type locality is PA. Although A. anna was described in the 1863 October–December issue of the Proceedings of the Entomological Society of Philadelphia (Volume 2), the title page to volume 2 indicates it was published in 1864, not 1863. The publication date of 1863 given for A. anna and A. persephone in Ferguson & Opler (2006) is therefore incorrect.
Arctia persephone : described from a male holotype, type no. 6886 in ANSP [not examined]; the type locality is PA.
Diagnosis: The broad black hindwing borders and lined forewings with prominent postmedial (and often medial) bands are usually sufficient to recognize G. anna . Rare forms of G. anna with reduced forewing bands may resemble large G. virguncula , but G. anna never has a thick-lined cubital vein, and vein Cu2 is continuous with or barely separated from the postcubital stripe in G. anna , whereas in G. virguncula Cu 2 remains well separated from the postcubital for its entire length. Yellow forms of G. virgo and G. parthenice lack the broad suffused hindwing markings characteristic of G. anna . Yellow forms of G. ornata can be separated by the lack of transverse band extensions beyond the postcubital stripe, and the fact that they occur on opposite sides of the continent in vastly different habitats.
Description: Head – Palps black, frons and vertex yellowish buff; male antennae moderately bipectinate, rami averaging 4.53 ¥ 10 - 1 mm, (N = 6); female antennae slightly biserrate; dorsal scales dark brown to black, varying to predominantly yellowish buff; eyes well developed. Thorax – Vestiture black with yellowish buff borders on vertex, patagia, and tegulae; patagia rarely entirely buff; entirely black ventrally, or with yellowish tuft at base of forecoxa; legs entirely or predominantly black, tibia yellowish laterally. Abdomen – Dorsal ground colour yellow, apex black; medial and lateral markings black; medial band relatively broad; pale buff ventrally, lateral black markings broad, often confluent such that ventrum is entirely black. Forewing – Male forewing length averaging 20.1 mm (N = 6 males); black dorsally, vein lines complete, lines and bands yellowish buff to whitish buff; antemedial band reduced to wide spot at costa or absent; medial band restricted to discal bar, sometimes reduced to line or absent entirely; postmedial band well developed, angled strongly at M 3, very rarely reduced or absent; subterminal band well developed; fringe and anal margin concolourous with pale markings, costal margin entirely pale, sometimes only to subterminal band; ventral markings similar, but dark markings with a paler yellowish cast; sexes similar. Hindwing – Ground colour yellow, black markings highly variable in extent; antemedial spots when present streaked basad; medial spot usually large and prominent; postmedial and subterminal elements usually confluent into one broad band, postmedials virtually never discrete; ventral markings similar, but dark markings with a paler yellowish cast; sexes similar, although females generally with more extensive dark markings, sometimes black entirely. Male genitalia – Distal portion of valve gradually tapering to rounded apex; clasper poorly developed, median ridge reduced; uncus broad-based, process evenly tapered to point, three ¥ as long as width of base; juxta 1.5–2¥ wider than height; aedeagus with dorsad curve at 2/3 distance beyond base; vesica with basal and medial chamber approximately equal in length and width, not scobinate; second medial diverticulum elongate, prominent; distal chamber relatively large, 2.5 ¥ as long as width of medial chamber, kidney-shaped, minutely scobinate; vesica as long as or slightly shorter than aedeagus. Female genitalia – Not examined.
Biology: Adult collection dates indicate a single flight period between late May and late June. Grammia anna inhabits relatively rich, mesic wooded areas, and is one of only a few species restricted to habitats associated with deciduous forests of eastern North America.
Distribution: Southern ON and New England south to GA. Smith (1938a) records it from as far west as eastern NE ( Fig. 102 View Figures 102–107 ).
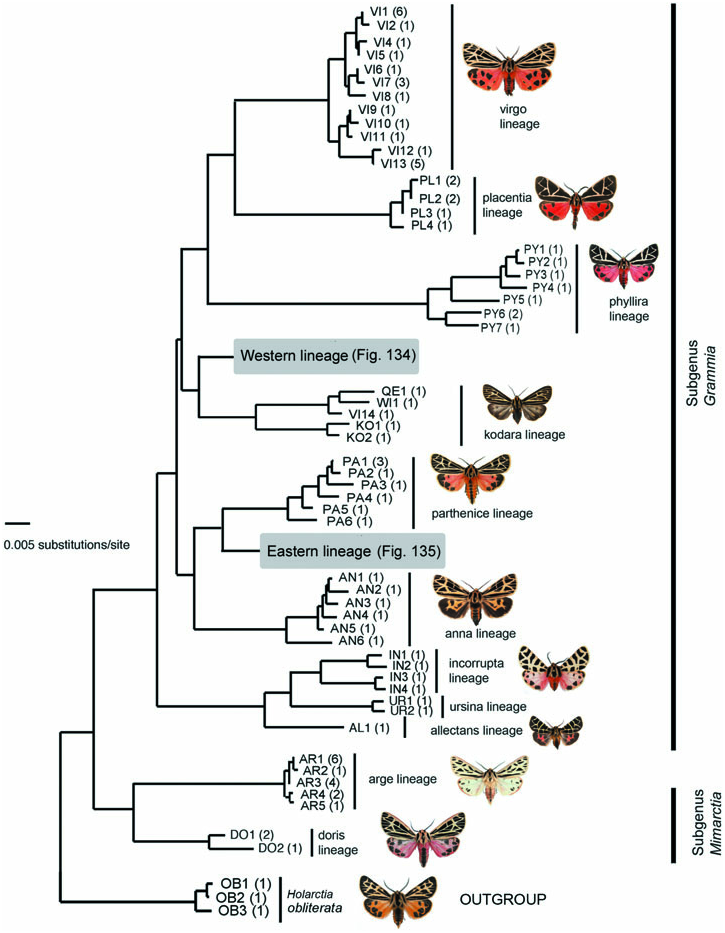
Molecular variation: Six specimens of G. anna from four localities (ON, QC, GA, NC) all exhibited unique haplotypes, at least 4.9% divergent from all other Grammia haplotypes ( Table 2).
GRAMMIA PARTHENICE (KIRBY) ( FIGS 10, 47, 78, 103)
Callimorpha parthenice Kirby, 1837: 204 .
Arctia saundersii Grote, 1864c: 75 , pl. 4 f. 3.
Arctia intermedia Stretch, 1873 [1874]: 216, pl. 9 f. 3. Arctia stretchii Grote, 1875: 197 .
Arctia approximata Stretch, 1885: 104 .
Arctia intermedia var. circa Stretch, 1906: 120 , pl. 5 f. 26.
Apantesis parthenice (Kirby) ; Franclemont, 1983: 117. Apantesis parthenice intermedia (Stretch) ; Franclemont, 1983: 117.
Grammia parthenice (Kirby) ; Ferguson & Opler, 2006: 9.
Grammia parthenice intermedia (Stretch) ; Ferguson & Opler, 2006: 9.
Type material: Callimorpha parthenice : described from an unstated number of types from ‘North America’; as there are no known extant types of taxa described by Kirby (1837), the following neotype (Fig. 10A) is designated to ensure the stability of the name: ‘CAN [ ADA]: AB [ AB] Chauvin, 13 km N/ 52.810N 110.106W / 25-vii-04 C. Schmidt/ sand dune parkland’, ‘ NEOTYPE / Callimorpha / parthenice Kirby / B.C. Schmidt, 2009’ [ CNC]. A number of taxa described by Kirby (1837) were collected by the Franklin expedition in present-day SK and northern AB, Canada, and the neotype is therefore selected from the same region.
Arctia saundersii : described from an unstated number of specimens, a male syntype in ANSP (type no. 7688) [not examined]; the type material was collected in ‘ Canada West’ , present-day southern ON, Canada. Hampson (1901) lists five male specimens in the Grote collection from Renfrew Co., Canada. The type locality of A. saundersii is therefore restricted to Renfrew County, ON, Canada to ensure the stability of the name for this geographically variable species .
Arctia intermedia : described from an unstated number of specimens from TX [ USA]. The probable male holotype is in CAS, type no. 3245 [not examined] .
Arctia stretchii : described from TX [ USA]; the male holotype is in USNM, type no. 33682 according to Smith (1938a) [not examined]. Based on a variant of parthenice that exhibits a complete antemedial band .
Arctia approximata : described from a single female specimen from ‘Canada’ [ CAS, not examined]; The illustration in Stretch (1906, plate 8 fig. 10) is likely to be of the type. Although the vague reference to ‘Canada’ in the 1885 description could apply to much of present-day Canada, it is seems most likely that the type material of approximata originated from the relatively well-known territory of southern ON (‘ Canada West’ prior to confederation in 1867); in order to stabilize the name approximata , the type locality is hereby restricted to Ottawa, ON, Canada.
Arctia intermedia var. circa : named based on an indication to an illustration, with no given type locality or known extant types; circa is an unavailable infrasubspecific name. The illustration in the original description is diagnostic, and is a variant of G. parthenice .
Diagnosis: Most similar to G. virgo , but G. parthenice lacks any trace of the antemedial spots on the hindwing that are so prominent in virgo ; the cubital vein is thin-lined in G. parthenice , thick in G. virgo .
Description: Head – Palps black, frons and vertex yellowish buff; male antennae moderately bipectinate, rami averaging 4.03 ¥ 10 - 1 mm, (N = 6); female antennae slightly to strongly biserrate; dorsal scales dark brown to black; eyes well developed. Thorax – Vestiture black with pale to yellowish buff (rarely pinkish buff) borders on vertex, patagia, and tegulae; entirely black ventrally, rarely with buff scales at base of forecoxa; legs entirely or predominantly black, tibia rarely buff laterally. Abdomen – Dorsal ground colour yellow-orange to orange-pink, apex black, rarely buff; medial and lateral markings black; medial band varying from thin to broad; buff ventrally, lateral black markings varying from lateral rows of spots to largely or entirely confluent, often entirely black. Forewing – Male forewing length geographically variable, ranging from 16.5 to 27 mm, females averaging slightly larger; black dorsally, vein lines complete, lines and bands yellowish buff to whitish buff, rarely pink or pinkish buff; antemedial band usually reduced to wide spot at costa or absent, sometimes complete and extending to anal margin; medial band usually well developed and restricted to discal bar, sometimes extending to postcubital stripe or beyond; postmedial band well developed, angled slightly at M 3, rarely reduced or absent; subterminal band well developed; fringe and anal margin concolourous with pale markings, costal margin entirely or (rarely) partly pale; ventral markings similar, but dark markings slightly paler, pale areas with distinct yellow cast; sexes similar, but females with slightly more elongate wing shape. Hindwing – Ground colour orange-pink to deep pink, particularly in females; black markings variable in extent; antemedial spots never present; medial spot usually large and prominent; postmedial and subterminal elements varying from very reduced to broad and confluent, particularly at tornus; ventrally with paler dark markings and ground colour washed-out and yellowish; sexes similar. Male genitalia – Distal portion of valve gradually tapering to rounded apex; clasper poorly to moderately developed, median ridge moderately developed; uncus broad-based, process evenly tapered to point, two to three ¥ as long as width of base; juxta 1.5–2¥ wider than height; aedeagus with dorsad curve at 2/3 distance beyond base; vesica with basal and medial chamber approximately equal in length and width, not scobinate; distal chamber twice as long as width of medial chamber, relatively narrow, elongate; vesica slightly shorter than aedeagus. Female genitalia – Ductus bursae unsclerotized, slightly constricted medially; corpus bursae globose, three ¥ width of ostium bursae; signa round to slightly elliptical; signa finely scobinate; coil of appendix bursae evenly rounded; posterior apophysis 2.5 ¥ longer than papillae anales.
Biology: Grammia parthenice is univoltine, with adult collection dates ranging from late June to early October; northern and Rocky Mountain records are primarily from mid July to early August. Collection dates for the eastern USA, at least as far north as PA and MA, are for September, and into October for NC, GA and west to TX. Grammia parthenice inhabits a wide variety of ecoregions and habitats, from rich deciduous forests to the grasslands of the Great Plains, preferring mesic meadows, pastures, and edges of wetlands.
Distribution: The most widespread species of Grammia , occurring from central BC east to NF ( Morris, 1980), south to GA in the east, west to TX, UT, and MT ( Fig. 103 View Figures 102–107 ).
Molecular variation: Twenty specimens of G. parthenice from ten localities across the species’ range segregated into nine haplotypes of two highly divergent lineages, resulting in up to 3.5% intraspecific divergence ( Table 2). Haplotypes of the Parthenice lineage ( Fig. 133 View Figure 133 ) formed a monophyletic group, but haplotypes of the Eastern lineage ( Fig. 135) were shared with or very similar to those of other species in the Eastern lineage. Eastern lineage haplotypes probably represent introgression of nonconspecific mtDNA into G. parthenice populations ( Schmidt & Sperling, 2008). Haplotypes of the Parthenice lineage were restricted to eastern North America ( QC, KY, NC, GA), whereas Eastern haplotypes occurred throughout the northern portion of the species’ range.
Remarks: Although G. parthenice is superficially most similar to G. virgo , molecular data did not suggest a close relationship between these species. The numerous phenotypical and physiological differences also do not support a close relationship between G. parthenice and G. virgo . The phenotypic differences are given above in the diagnosis; the fact that G. virgo flies in mid summer whereas G. parthenice flies in late summer/early autumn is indicative of physiological differences. Shared haplotypes between G. parthenice and G. virguncula may indicate a closer relationship between these species, but they differ in forewing pattern (presence of transverse bands and finer vein lines in parthenice ), hindwing colour and pattern (pink-orange not yellow, never melanic; lacking basal dark markings) abdomen colour and pattern (pinkorange, last two segments sometimes entirely black), and the relatively shorter antennal rami (relative to body size), larger size and earlier flight period of parthenice . Grammia parthenice and G. virguncula occur sympatrically over most of eastern North America, and I have not seen phenotypically intermediate specimens which could be attributed to hybridism.
Grammia parthenice is one of the most variable members of the genus in terms of size, colour, pattern, and genitalic structure. Although there is some geographical structure to this variation, much of it is also exhibited within populations. Specimens from the south-eastern USA often have wingspans more than twice that of the northern plains and boreal populations, and have been referred to as subspecies intermedia ( Franclemont, 1983; Ferguson & Opler, 2006); this name was based on type material from TX, so it may not apply to the eastern and south-eastern populations. This geographical variation should be re-evaluated with more study material.
(FIGS 11A–C, 49, 79, 104)
Callimorpha virguncula Kirby, 1837: 304 .
Euprepia virguncula var. otiosa Neumögen & Dyar, 1893: 142 .
Apantesis virguncula (Kirby) ; Franclemont, 1983: 117.
Grammia virguncula (Kirby) ; Ferguson & Opler, 2006: 9.
Type material: Callimorpha virguncula: Described from an unstated number of specimens from ‘Canada’; Kirby’s type specimen(s) is presumed lost, as there are no known extant types of any of the taxa described by Kirby in 1837, including several butterfly species ( Miller & Brown, 1981) and two plusiine noctuids ( Lafontaine & Poole, 1991); in order to stabilize the nomenclature of this species complex, the following neotype (Fig. 11A) is designated: ‘ Canada, Ontario,/ Carp Ridge, E side/ 45 24.63’N 76 03.45′W / 28 June 2003 / J. Troubridge, leg.’, ‘ NEOTYPE / Callimorpha / virguncula Kirby / B.C. Schmidt, 2009’ [ CNC].
Euprepia virguncula var. otiosa : the name is based on two aberrant specimens of virguncula which show traces of transverse bands, and otiosa is therefore an unavailable infrasubspecific name.
Diagnosis: Grammia virguncula is most similar to G. speciosa , G. kodara , and G. philipiana . In comparison to G. speciosa , virguncula has a thick, not thin-lined cubital vein, has a more truncate/broad forewing shape, is larger overall, and has a thin, not thicklined cubital vein; the hindwing black markings are also usually less developed in virguncula . Also, G. virguncula is a nocturnal species that comes to light, whereas speciosa is diurnal, reflected in the smaller eye size of speciosa . Grammia virguncula and G. kodara do not occur together, and the differences between these taxa are discussed under G. kodara . Compared to G. philipiana , G. virguncula has yellow hindwings, not orange, longer male antennal branches, and normal, not reduced, eyes. Holarctia obliterata is superficially similar, but has better developed forewing costal bars, in addition to the substantial genitalic differences discussed under that species and the discussion of the genus Holarctia .
Description: Head – Palps black, frons and vertex pale to yellowish buff, vertex rarely black centrally; male antennae moderately bipectinate, rami averaging 4.35 ¥ 10 - 1 mm, (N = 19 males); female antennae moderately to strongly biserrate; dorsal scales dark brown to black; eyes well developed, mean diameter 8.68 ¥ 10 - 1 mm. Thorax – Vestiture black with pale to yellowish buff borders on vertex, patagia, and tegulae; black ventrally, base of coxa and distal area usually with yellowish buff scales; coxa and femur black centrally, yellow at base and apex, giving striped appearance; tibia and tarsus pale buff dorsally, black ventrally. Abdomen – Dorsal ground colour yellow, pale buff near apex; medial and lateral markings black; pale buff ventrally, lateral black markings broad but not confluent at midline. Forewing – Male forewing length averaging 16.2 mm (range 13.9– 20.5 mm, N = 15 males), largest for populations from the central Missouri River region (SD, NE, IA, KS); black dorsally, vein lines complete, lines and bands yellowish buff to pale whitish buff; cubital vein line often broader than other vein lines; all bands except subterminal usually absent, medial band very rarely present as bar in discal cell; subterminal band complete; fringe and anal margin concolourous with pale markings, costal margin entirely pale, sometimes reduced to basal half; ventral markings similar, but dark markings with a paler yellowish cast; sexes similar. Hindwing – Ground colour yellow, black markings highly variable in extent, but antemedial, medial, postmedial, and subterminal elements nearly always present; antemedial elements usually as streaks or dashes extending to base, submarginal spots often discrete; ventral markings similar, but dark markings with a paler yellowish cast; sexes similar. Male genitalia – Distal portion of valve gradually tapering to rounded or slightly pointed apex; clasper poorly to moderately developed, median ridge moderately developed; uncus broad-based, process evenly tapered to point, two to three ¥ as long as width of base; juxta 1.5–2¥ wider than height; aedeagus with dorsad curve at 2/3 distance beyond base; vesica with basal and medial chamber approximately equal in length and width, not scobinate; distal chamber medium-sized, twice as long as width of medial chamber, kidney-shaped, roughly scobinate; vesica as long as or slightly shorter than aedeagus. Female genitalia – Ductus bursae unsclerotized; proximal half of ductus bursae twice as wide as distal half; corpus bursae more or less globose, four¥ width of ostium bursae; signa round to slightly elliptical, relatively small averaging about 1.97 ¥ 10 - 1 mm; signa coarsely scobinate; coil of appendix bursae evenly rounded; posterior apophysis 1.5–2¥ longer than papillae anales.
Biology: The peak flight period is from mid June to late July, with extreme dates spanning late May to late August. Grammia virguncula occurs in a variety of open wooded habitats, ranging from marshes, fens, and bogs to transition parkland and prairie. Despite this apparent range of suitable habitats, virguncula is usually not common. Adults come to light.
Distribution: Occurs across most of southern Canada and the USA from the Rocky Mountains eastward. Virguncula reaches its northern range limit in the boreal forest (northern AB and NF), and occurs south along the Rocky Mountain system to Apache Co., AZ, and NJ in the east.
Molecular variation: Grammia virguncula exhibited 19 haplotypes among 47 sampled individuals, from 20 localities across most of the species’ range ( Table 2). Much of the Eastern lineage consisted of G. virguncula haplotypes, and only one G. virguncula haplotype fell outside this lineage ( VR 1, Western lineage: Fig. 134 View Figure 134 ). Thirteen of 19 haplotypes were unique to G. virguncula , although most of these differed by only one or two base pairs from haplotypes occurring in other species ( Figs 134 View Figure 134 , 135). Species sharing haplotypes with virguncula included G. williamsii , G. parthenice , G. figurata , and G. margo , all within the Eastern lineage. The Eastern lineage is most likely native to G. virguncula , with multiple hybridization events resulting in taxonomically extensive introgression of Eastern haplotypes into other species ( Schmidt & Sperling, 2008). Despite extensive haplotype sharing, there is genetic evidence that the phenotypically distinct Atlantic maritime populations are unique ( Schmidt, 2007). Similarly, no haplotypes were shared with the closely related G. speciosa ( Fig. 135). As a result of the complex genealogy, the currently available molecular data are insufficient to resolve any potential genetic differentiation of virguncula subspecies discussed below.
Remarks: Specimens from the Missouri River basin between south-eastern SD and western MO are larger with more reduced hindwing dark markings (Fig. 11E) compared to those from the adjacent Great Lakes and Great Plains regions and should perhaps be considered as a distinct subspecies, but more study material is needed.


| ANSP |
Academy of Natural Sciences of Philadelphia |
| QC |
National Museum of Natural History, Bulawayo |
| ADA |
Department of Agriculture |
| CNC |
Canadian National Collection of Insects, Arachnids, and Nematodes |
| CAS |
California Academy of Sciences |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| UT |
University of Tehran |
| MT |
Mus. Tinro, Vladyvostok |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Grammia anna
| Schmidt, B. Christian 2009 |
Grammia parthenice (Kirby)
| Ferguson DC & Opler PA 2006: 9 |
Grammia parthenice intermedia (Stretch)
| Ferguson DC & Opler PA 2006: 9 |
Grammia virguncula (Kirby)
| Ferguson DC & Opler PA 2006: 9 |
Apantesis anna (Grote)
| Ferguson DC & Opler PA 2006: 9 |
| Franclemont JG 1983: 117 |
Apantesis parthenice (Kirby)
| Franclemont JG 1983: 117 |
| Franclemont JG 1983: 117 |
Apantesis virguncula (Kirby)
| Franclemont JG 1983: 117 |
Arctia intermedia var. circa
| Stretch RH 1906: 120 |
Euprepia virguncula var. otiosa Neumögen & Dyar, 1893: 142
| Neumogen B & Dyar HG 1893: 142 |
Arctia approximata
| Stretch RH 1885: 104 |
Arctia intermedia
| Grote AR 1875: 197 |
Arctia anna
| Grote AR 1864: 335 |
Arctia persephone
| Grote AR 1864: 433 |
Arctia saundersii
| Grote AR 1864: 75 |
Callimorpha parthenice
| Kirby WF 1837: 204 |
Callimorpha virguncula
| Kirby WF 1837: 304 |