
Glabrilaria hirsuta Rosso, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4524.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:22A47EBE-338A-4AAF-A63F-45EDE9727F18 |
|
DOI |
https://doi.org/10.5281/zenodo.6485592 |
|
persistent identifier |
https://treatment.plazi.org/id/03BA0525-3A63-FFA6-5CAA-7FF4767CD449 |
|
treatment provided by |
Plazi |
|
scientific name |
Glabrilaria hirsuta Rosso |
| status |
sp. nov. |
Glabrilaria hirsuta Rosso n. sp.
( Figs 10–38 View FIGURES 10, 11 View FIGURES 12–18 View FIGURES 19–25 View FIGURES 26–28 View FIGURES 29–38 ; Tables 1, 2)
Material examined. Holotype: Great Bahama Bank slope, R/ V Maria S. Merian Cruise MSM20 View Materials /4, Station GeoB16374-1 one live, fertile colony encrusting a coral branch and including the ancestrula)—SMF-45.505 . Paratypes: Great Bahama Bank Slope, R / V Maria S. Merian Cruise MSM20 View Materials /4. Station GeoB16367-2 (two live and 20 dead colonies, all on corals, except for three on erect bryozoans, one on a serpulid, and one on a barnacle plate) – SMF-45.506; Station GeoB16368-1 (one dead colony on a coral)—SMF-45.507; Station GeoB16375-1 (31 dead colonies on corals)—SMF-45.508; Station GeoB16376-1 (3 dead colonies on corals)—SMF-45.509; Station GeoB16377-1 (one living and 12 dead colonies on corals, except for one on a fragment of sponge skeleton)—SMF- 45.510; Station GeoB16378-1 (one dead colony on a coral branch)—SMF-45.511; Station GeoB16382-1 (nine living and 30 dead colonies on corals)—SMF-45.512; Station GeoB16388-3 (one living plus two dead colonies on corals)—SMF-45.513 .
A total of six paratypes, four from Station GeoB16375-1, one from Station GeoB16377-1, and one live colony on a coral from Station GeoB16388-3: Rosso-Bahama Collection Bah.H., PMC B 21. 30.6.2017.
Etymology. From the Latin, hirsutus, alluding to the prickly, thorny appearance of the frontal shield and the ooecium.
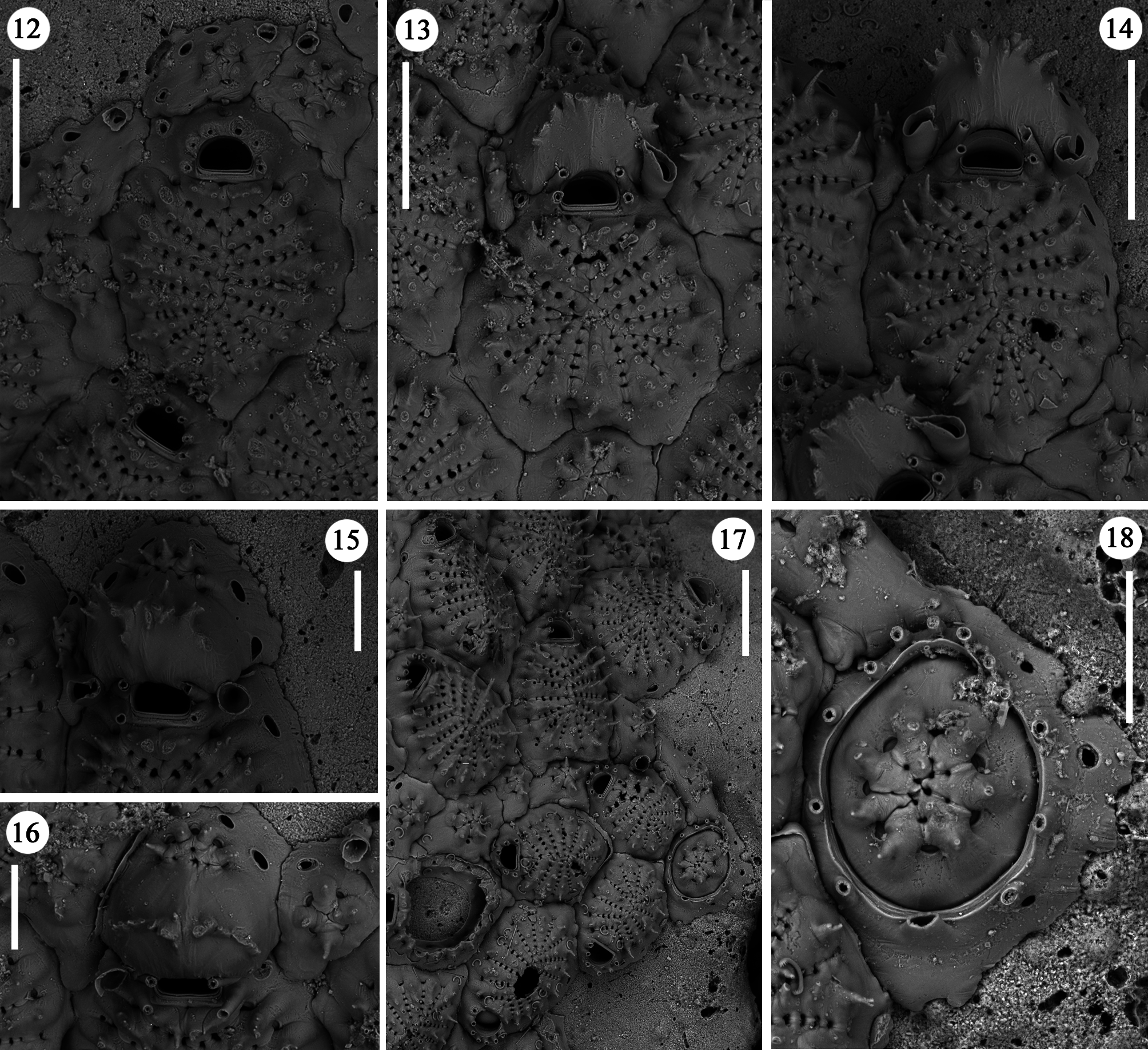
Description. Colony encrusting unilaminar, pauciserial, small-sized, usually including few tens of zooids, sometimes more, irregularly elongated to ribbon-shaped ( Fig. 10 View FIGURES 10, 11 ), resulting from a single lobe or two lobes starting from the ancestrula and growing usually in opposite directions, sometimes bifurcating; lobes usually less than 1 mm wide and 1–5 mm long.
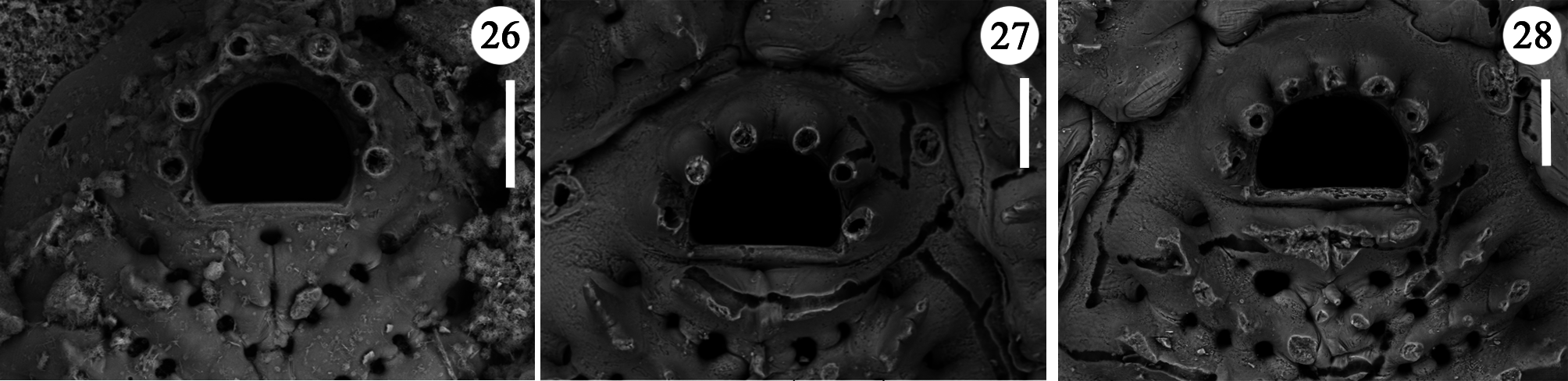
Zooids oval to roughly polygonal, small, longer than wide, moderately convex, distinct, the boundaries marked by deep furrows ( Figs 12–14 View FIGURES 12–18 , 21 View FIGURES 19–25 , 30–31 View FIGURES 29–38 ); outlines irregularly sinuate. Gymnocyst narrow, deeply sloping towards the basal surface, 10–40 µm wide, but occasionally forming prominent tips, wedging between adjacent zooids, irrespective of their position ( Figs 22, 24 View FIGURES 19–25 ). Communication in autozooids through c. 10 transversely oval windows of moderate size, positioned at the centre-line of distolateral walls ( Figs 12, 14, 15 View FIGURES 12–18 ), c. six in kenozooids. Frontal shield subcircular to oval, occupying almost the entire frontal surface, formed by 14–18, but usually 16–17 wedgeshaped costae, converging towards the centre in the circular shields ( Fig. 13 View FIGURES 12–18 ) or towards a median slightly raised carina in the oval shields ( Figs 11 View FIGURES 10, 11 , 15 View FIGURES 12–18 ). Costae joined by 5–6 bridges leaving regularly spaced, oval to kidneyshaped intercostal pores about 8–10 µm wide ( Fig. 25 View FIGURES 19–25 ), often bearing denticles, which mark the fusion of lateral processes. Only 3–4 intercostal pores are present proximally to the suboral pair of triangular costae that outline an almost right angle ( Figs 26–28 View FIGURES 26–28 ). Suboral pair of costae stouter, merging at the midline and often forming a transversal crest of pointed, upward directed, spine-like processes nearly orthogonal to the frontal surface, and leaving two small round pores in the midline, one just proximal to the orifice and another on the opposite side of the transversal crest ( Figs 14 View FIGURES 12–18 , 27, 28 View FIGURES 26–28 ). Roundish papilla pores are seen between bases of costae, inside the peripheral intercostal pores. Costae with a median carina progressively raising towards zooidal margins, bearing up to four prominent, usually simple but sometimes bifid and even trifid, spine-like processes, the marginal ones, the longest, giving a concave appearance to the shield in heavily spinous zooids ( Figs 11 View FIGURES 10, 11 , 14 View FIGURES 12–18 , 25 View FIGURES 19–25 ). Spiny processes also mark the shield midline carina, when present ( Figs 19 View FIGURES 19–25 , 29, 30, 38 View FIGURES 29–38 ). Orifice transversally D-shaped ( Figs 12 View FIGURES 12–18 , 20 View FIGURES 19–25 , 26– 28 View FIGURES 26–28 ), with the straight proximal border marked by an elevated rim and a curved distal one, slightly wider than long, provided with six articulated oral spines, seven on some periancestrular zooids and rarely also on adult zooids ( Fig. 28 View FIGURES 26–28 ), long (50 µm) and slender, articulated on equally spaced bases, sometimes coalescing and forming a palisade distal to the orifice ( Fig. 36 View FIGURES 29–38 ); four spines persisting on ovicellate zooids ( Figs 13–16 View FIGURES 12–18 , 20 View FIGURES 19–25 ), the distal ones abutting the proximal ooecium and slightly converging.
Ovicells hyperstomial, cleithral. Ooecium formed by the distal kenozooid [type 1 sensu Ostrovsky (1998, 2008, 2013) and type B sensu Bishop & Househam (1987)], with 3–4 spiny costae, visible in frontal view ( Figs 13, 15, 16 View FIGURES 12–18 , 35 View FIGURES 29–38 ). Ectooecium with a usually barely visible longitudinal suture, but characterised by a planar semicircular to sub-triangular proximal area steeply inclined, bordered by aligned, pointed, spine-like processes directed distally and upwards, the central ones the most prominent, forming a transversal crest ( Figs 13–16 View FIGURES 12–18 , 19– 21 View FIGURES 19–25 ). A second crown of 3–6 spiny processes is situated distally, borne on the costate frontal shield of the kenozooid ( Figs 15, 16 View FIGURES 12–18 ).
Adventitious avicularia pedunculate, their cystid erect, from cone-shaped to elongated bell-shaped, with more or less narrow base, and variably directed, triangular rostrum, steeply inclined to the colony surface, and lacking pivotal denticles ( Figs 36, 37 View FIGURES 29–38 ). Mandible triangular ( Fig. 37 View FIGURES 29–38 ). Avicularia sparse, originating from basal porechambers on both autozooids and kenozooids (irrespective of the presence of an ovicell), often in clusters of 2–3 from adjacent pore-chambers ( Figs 35–37 View FIGURES 29–38 ), easily detachable, and frequently lost, even in living colonies.
Kenozooids very numerous, often more numerous than autozooids in certain colonies or colony portions, clustered together ( Fig. 29 View FIGURES 29–38 ) or forming single ( Figs 12, 17 View FIGURES 12–18 , 21 View FIGURES 19–25 , 30 View FIGURES 29–38 ) or double rows between autozooids, and commonly located along colony margins ( Fig. 10 View FIGURES 10, 11 ); more rarely interspersed between zooids ( Fig. 38 View FIGURES 29–38 ). They are circular to polygonal in shape, often irregular, and, although variably sized, usually small in comparison to autozooids and less elevated. Kenozooids have extensive gymnocyst and a cribrimorph frontal shield of 4–7, usually six, costae in a radial pattern ( Figs 12, 17 View FIGURES 12–18 , 19, 21 View FIGURES 19–25 , 29–35 View FIGURES 29–38 ); each costa bearing 1–2, sometimes three pointed, often very prominent spine-like processes ( Fig. 32 View FIGURES 29–38 ). Large kenozooids with up to 10 costae have been occasionally found ( Fig. 31 View FIGURES 29–38 ). Particularly small (less than 100 µm in maximum length), and irregularly shaped kenozooids with reduced, ill-defined or even lacking costate shield occur on broken zooids or are aligned in depressions between zooids in damaged colony portions ( Figs. 33, 34 View FIGURES 29–38 ), clearly forming reparative patches.
Ancestrula tatiform ( Figs 17, 18 View FIGURES 12–18 , 22–24 View FIGURES 19–25 ) bearing the bases of an unpaired proximal spine, and 5–6 pairs of spines borne around the edge of the gymnocyst; the two distal pairs around the opesia more closely spaced than the others. Ancestrula occupied by an intramural reparative kenozooid in all cases observed. The first zooid is budded from the ancestrula distolaterally; further budding typically continues just on one side of the ancestrula, and its lateral walls remain partly exposed ( Figs 17 View FIGURES 12–18 , 22 View FIGURES 19–25 ). Only one ancestrula was found being completely surrounded by zooids ( Fig. 24 View FIGURES 19–25 ).
Remarks. Glabrilaria hirsuta Rosso n. sp. significantly differs from other cribrimorphs and clearly falls within Glabrilaria as defined above. It is characterised by the small size of colonies and zooids, colony morphology, heavy spiny appearance of the colony surface, abundance and clustering of kenozooids, and ooecia with a transverse series of prominent processes. However, spine-like processes may develop at variable extent on zooids and especially on ooecia in different colonies. In some colonies and/or colony portions, ooecia even show only two very prominent median spiny processes. The ooecial outline could also vary from round ( Figs 13–16 View FIGURES 12–18 ) or roughly triangular, about as long as wide, to irregularly quadrangular, wider than long ( Fig. 20 View FIGURES 19–25 ). Ooecia may or may not be associated with stalked avicularia that are usually paired, and could be located near their proximal sides, borne on the maternal zooid ( Figs 13–16 View FIGURES 12–18 ), or on their distal sides, borne on the associated kenozooid ( Fig. 21 View FIGURES 19–25 ). Stalked avicularia may be apparently lacking in some colonies owing to their detachment, mostly in the central, oldest colony portions, and are usually particularly evident at colony margins because these areas correspond to the young, recently grown portions, and because there the avicularia stand up against the substratum background. Spiny processes and stalked avicularia may be hardly detectable on worn specimens, although they are extremely distinctive in live and well-preserved colonies. Intramural reparative regeneration was also very common in the studied material. Particular breakage of the frontal shields was also detected ( Figs 13, 14, 17 View FIGURES 12–18 , 33 View FIGURES 29–38 ) that can be, at least partly, interpreted as evidence of predation. In some instances, such broken zones are covered with small patches of frontal calcification ( Fig. 34 View FIGURES 29–38 , arrowed). Also several orifices show inner (double and even third) rims ( Figs 12–16 View FIGURES 12–18 ), pointing to the regeneration of certain zooids following predation (Berning 2008). These evidences indicate the heavy predation pressure on this species, and its difficult endeavour to protect itself notwithstanding the large number of oral spines and frontal spinous processes.
Periancestrular astogenetic pattern is distinctive and leads to the development of elongated colonies where the ancestrula is located at one end of a single lobe or in between two ‘lateral’ lobes. In contrast, the ancestrula’s proximal budding loci seems to become active only late in astogeny. Several colonies in the present material were young and included a small number of zooids. Lobate pauciserial colonies are not uncommon in deep settings, although lobes tend to be fan-shaped rather than ribbon-like, and to develop distally to the ancestrula as in Herentia hyndmanni (Berning et al. 2008) and Atlantisina species ( Berning et al. 2017), among others. Within cribrilinids, Cribrilaria arrecta ( Bishop & Househam, 1987) n. comb. forms lobes with roughly unchanged width, which are, however, wider than in the present species. Cribrilaria arrecta n. comb. also shares with G. hirsuta Rosso n. sp. prominent carinate costae with pointed processes, but it significantly differs in several characters, among others in the large interzooidal avicularia with flabellate mandibles, a lower number of costae, and a different suboral bar.
The zooidal spiny appearance of G. hirsuta Rosso n. sp. is reminiscent of specimens from the Kermadec region recorded as Puellina (Cribrilaria) innominata (Couch) by Gordon (1984, pl. 21, fig. C), but other features, including the number of costae and oral spines, as well as the type of ovicell and avicularia, are completely different.
Glabrilaria hirsuta Rosso n. sp. is clearly distinct from most species now allocated in Glabrilaria View in CoL (see above), but resembles both G. pedunculata View in CoL and G. cristata View in CoL in its spinous colony surface, the common kenozooids, and the erect columnar avicularia. However, G. pedunculata View in CoL has, among other characters, spot-like colonies, a relatively smooth appearance, and an ooecium with a prominent longitudinal carina (Harmelin 1988, figs 10, 11; Rosso et al. 2018, fig. 5e, f). On the other hand, as stated by Harmelin (1978: 187), and as it is evident from his images (fig. 7 and pl. 1, figs 8–10), G. cristata View in CoL has more convex zooids with a shield of less numerous, yet more thorny, costae (ranging from a mean of 9.5 to 13 in colonies collected at 300 and 600 m depth, respectively), each bearing seven cristate spine-like processes converging in a sensibly raised carina; a suboral shelf including several intercostal spaces and only occasionally a single one; 4–5 costae in kenozooidal frontal shield; seven oral spines; a raised carina on the ooecium besides the spiny transversal one; and shorter zooids. However, the most distinctive character of G. cristata View in CoL is the “peristomial” structure formed by the first pair of costae that elevate in a distally incomplete, denticulate collar that partly encircles oral spines. This structure is completely missing in G. hirsuta Rosso n. sp.
Both G. cristata View in CoL and G. hirsuta Rosso n. sp. occur in the shallow bathyal zone between 135–900 m ( Harmelin 1978) and 572–660 m depth interval (present data), respectively, and from temperate to subtropical latitudes at 29– 39° and 24–25° N, respectively. While G. hirsuta Rosso n. sp. is restricted to the western Atlantic, G. cristata View in CoL occurs in central Atlantic seamounts and the Azores Archipelago. Glabilaria cristata View in CoL is associated with the deepwater corals Madrepora oculata View in CoL and Desmophyllum dianthus View in CoL (two scleractinian coral species also reported from the Western Atlantic: see above), whereas G. hirsuta Rosso n. sp. was found preferentially on fragments of the dendrophylliid Enallopsammia profunda View in CoL from coral rubble-rich sediments. In both Glabrilaria View in CoL species the development of spiny processes on the costae and spiny projecting processes associated with the orifice and the ooecia, although differently shaped, could represent an adaptation for trying to cope with predation pressure in their habitats (see above). Both species are known exclusively based on present-day living populations and isolated dead specimens.
Distribution. Glabrilaria hirsuta Rosso n. sp. is presently known only from the GBBS, with live colonies found at 572–660 m depth (see above), and dead specimens extending down to 677 m. Colonies were mostly found on erect branched corals, but encrusted also skeletons of other benthic invertebrates, including erect bryozoans, barnacle plates and the spicule net of the sponge Aphrocallistes . The finding of live specimens at station GeoB16388, where the occurrence of seepages is pointed out by lucinid associations ( Hebbeln et al. 2012), could indicate a certain environmental tolerance of this species.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Ascophorina |
|
Family |
|
|
Genus |
Glabrilaria hirsuta Rosso
| Rosso, A., Beuck, L., Vertino, A., Sanfilippo, R. & Freiwald, A. 2018 |
Glabrilaria hirsuta Rosso
| Rosso & Beuck & Vertino & Sanfilippo & Freiwald 2018 |
G. hirsuta Rosso
| Rosso & Beuck & Vertino & Sanfilippo & Freiwald 2018 |
G. hirsuta Rosso
| Rosso & Beuck & Vertino & Sanfilippo & Freiwald 2018 |
G. hirsuta Rosso
| Rosso & Beuck & Vertino & Sanfilippo & Freiwald 2018 |
G. hirsuta Rosso
| Rosso & Beuck & Vertino & Sanfilippo & Freiwald 2018 |
Glabrilaria
| Bishop & Househam 1987 |
Glabrilaria
| Bishop & Househam 1987 |
G. pedunculata
| Gautier 1956 |
G. pedunculata
| Gautier 1956 |
Madrepora oculata
| Linnaeus 1758 |