
Fridericia longeaurita, Boros, Gergely & Dózsa-Farkas, Klára, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3911.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:744769AD-E4D6-4AD3-A067-4006095378F5 |
|
DOI |
https://doi.org/10.5281/zenodo.5679665 |
|
persistent identifier |
https://treatment.plazi.org/id/03B3B403-FF99-CD37-FF26-755A616A946E |
|
treatment provided by |
Plazi |
|
scientific name |
Fridericia longeaurita |
| status |
sp. nov. |
Fridericia longeaurita View in CoL sp. n.
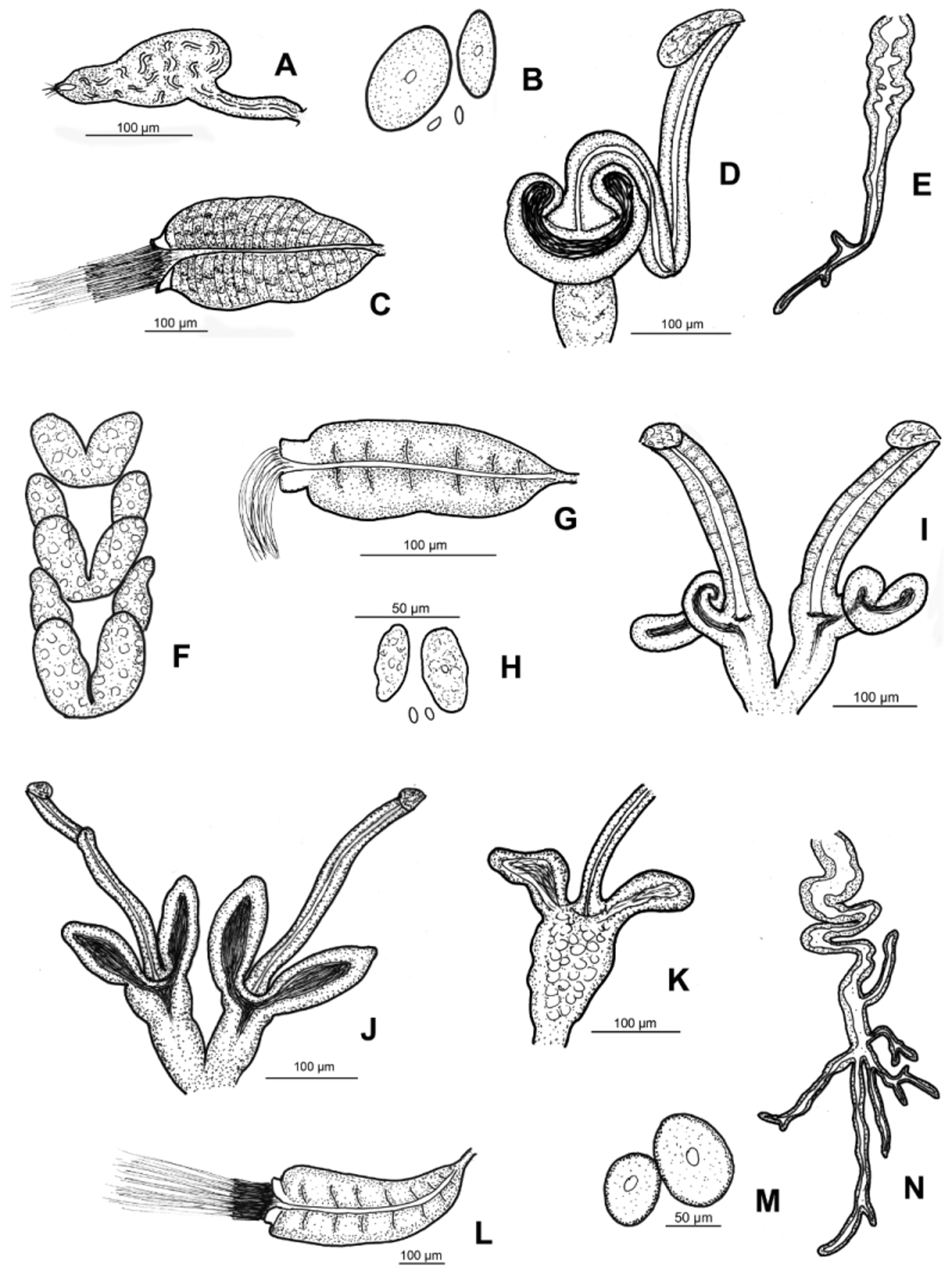
( Figs 2 View FIGURE 2 J–N, 6–7)
Type material. Holotype: F.21, slide No. 1024. Type locality: N46.14613 E24.63541, 440 m asl, close to Malancrav village, Romania. Leg. G. Boros, 15.10.2013. Adult specimen whole-mounted in euparal without staining. Paratypes: P.98.1–5, slides No. 1013, 1014, 1022, 1036, 1061, 5 adult whole mounts. N46.14613 E24.63541, 440 m asl, close to Malancrav village, Romania. Leg.: G. Boros, 15. 10. 2013. P.98.6–7, slides No. 1021, 1029, 2 adults, whole mounts. N46.13445 E25.18995, 590 m asl, hills over the village of Beia, Romania. Leg. G. Boros, 15.10.2013. (Slides No. 1013, 1014, 1021 and 1022 stained with borax-carmine; slides No. 1029 and 1061 stained with bromophenol blue; slide No. 1036 stained first with bromophenol blue, later with boraxcarmine.)
Etymology. Named after the shape of spermathecal diverticula which are mostly elongate (=longus) and earshaped (=aurita).
Diagnosis. The new species can be recognized by the following combination of characters: (1) large size (14–26 mm in vivo), segments 38–67; (2) maximum of 4 (5) chaetae per bundles preclitellarly, but only two or one postclitellarly; (3) clitellum girdle-shaped, weakly developed: hyalocytes and granulocytes arranged in transverse rows; (4) five pairs preclitellar nephridia; (5) coelomo-mucocytes large, with small refractile granules at periphery, lenticytes large; (6) chylus cells in XIII–XVI; (7) large seminal vesicle; (8) subneural glands absent; (9) sperm funnel cylindrical, approximately 3/4 as long as body diameter, collar narrower than funnel body, sperm heads 80–90 µm long in vivo; (10) spermathecal ectal duct long, but shorter than body diameter, one small sessile ectal gland at the orifice; ampullae with two long sac-like diverticula, one common opening dorsally into oesophagus.
Description. Holotype 25 mm long, 500 µm wide at VIII and 570 µm at clitellum in vivo (19.7 mm long, 540 µm wide at VIII and 590 µm at the clitellum, fixed), 67 segments. Body length of paratypes 14–26 mm, width on average 490±40.7 µm at VIII and 530±52.6 µm at clitellum (in vivo), length of fixed specimens 11–20 mm, width on average 513±84 µm at VIII, 542±65 µm at clitellum, segments 38–67. Chaetal formula: 2,3,4 – 2,1: 2,3,4,(5) – 2,(3,1). (In one bundle in VI of 1 specimen 6 chaetae, slide No. 1029). In contrast to most Fridericia species, chaetae within a bundle of same length (about 40–60 µm) and thickness (4–5 µm), sometimes one chaeta in a bundle slightly shorter and thinner than the others (35– 40 x 3–4 µm) ( Fig. 6 View FIGURE 6 C). At the posterior body end chaetae about as long as preclitellarly. Head pore at 0/I. Dorsal pores from VII. Epidermal gland cells arranged in 2–3 transverse rows per segment. Clitellum in XII–1 /2XIII, girdle-shaped, hyalocytes and granulocytes arranged in transverse rows, weakly developed especially ventrally. Thickness of body wall about 30–45 µm, cuticle about 1–1.5 µm in fixed specimens. Brain egg-shaped, about 160–190 Μm long and about 1.8 times longer than wide (fixed) ( Fig. 6 View FIGURE 6 A). Septa 6/7–9/10 thickened ( Fig. 6 View FIGURE 6 B).
Oesophageal appendages in IV winding, long, extending to VI, with several long and short terminal branches ( Fig. 2 View FIGURE 2 N). Pharyngeal glands all united dorsally and with ventral lobes. Chloragocytes from V, about 21–28 Μm long. Dorsal vessel from XIX–XXII, blood colourless. Midgut pars tumida in XXXI–XLV, occupying 5–13 (mostly 8–10) segments (in one case only three segments). Five pairs of preclitellar nephridia from 6/7 to 10/11, length ratio anteseptale: postseptale 1: 1.3–1.8, midventral origin of efferent duct ( Fig. 6 View FIGURE 6 D). Coelomocytes light brown in vivo, filling the coelom. Coelomo-mucocytes large, length 30–77 Μm in vivo, 25–48 µm fixed, oval (e.g. 47 x 35 Μm or 66 x 46 µm) and with a very characteristic and clear outline marked by fine, refractile granules at periphery ( Figs 2 View FIGURE 2 M, 6E, F,); granules very small, all of same size, only at periphery ( Fig. 6 View FIGURE 6 F). Lenticytes large 9–13 Μm long (in vivo). Chylus cells between XIII–XVI, occupying 3–4 segments ( Fig. 6 View FIGURE 6 G). Seminal vesicle large in XI–XII or XII–XIII. Sperm funnels ( Figs 2 View FIGURE 2 L, 6H. I) cylindrical, on average 330±85 µm long and 2–2.8 times as long as wide (in vivo); funnel length in fixed specimens on average 243±62 µm. Collar distinctly narrower than funnel body (about 89 µm) and relatively high (30–40 µm) in vivo. Length of spermatozoa not measurable, heads on average 69±9 µm in fixed specimens. Diameter of sperm ducts 9–12 µm (in vivo). Male copulatory organs 140–170 µm long, 90–110 µm wide and 50–66 Μm high (in vivo), bursal slits roughly H-shaped but mostly one branch somewhat longer than the others ( Fig. 7 View FIGURE 7 A). Subneural glands absent. Spermathecae ( Figs 2 View FIGURE 2 J, K, 7B–F): one small sessile ectal gland at the orifice, length about 22–35 µm (in vivo). Ectal ducts on average 332±62 µm long and 22±3,8 µm in vivo (302±58 µm long when fixed), canals not widened. Ampullae with two sac-like diverticula, often tapering distally, mostly sperm in lumina ( Fig. 7 View FIGURE 7 B–F). Length of diverticula 73–138 µm, diameter 40–60 µm in vivo. Ampullar proximal part mostly set off from distal part, fused with one common opening into oesophagus. Joint opening was not always distinguished due to overlaying pharyngeal glands. One mature egg at a time.
Distribution and habitat. Romania: Transylvanian Plateau, close to Malancrav and Beia villages. Shrub pasture, meadows.
Differential diagnosis. The new species is most similar to F. w a l d en s t ro e m i Rota & Healy, 1999 on the basis of shape of spermataecae but clearly separate from this species in the following main respects: in F. waldenstroemi the seminal vesicle is absent, chylus cells occur preclitellarly, the dorsal vessel originates in XIV–XVIII and the oesophageal appendage is of the a-type, i.e. short and unbranched ( Rota & Healy 1999), while the new species has a large seminal vesicle, chylus in XIII–XVI, dorsal vessel from XIX–XXII, and the oesophageal appendage is large with more branches at the end piece. F. montafonensis Schmelz, 1998 is also large (13–16 mm and 49–71 segments), has slightly similar spermathecae and also a large seminal vesicle, but the spermathecal diverticula are shorter, there is a large spermathecal ental bulb, and the two diverticula form a common U-shaped spermcontaining chamber together with the distal part of the ampulla. The coelomo-mucocytes are of the c-type, the chylus cells are located farther back (in XIV–XIX), the sperm funnel is shorter (100–170 µm long) and with collar as wide as the funnel ( Schmelz 1998, 2003). Furthermore, F. montafonensis has large subneural glands in XIII and XIV. F. brachiata Rota, 1994 is also large (10–22 mm long with 47–63 segments) and the spermathecae have long diverticula, but the spermathecal ectal ducts are shorter (only about 100 µm in contrast to about 300 µm in the new species), the coelomo-mucocytes are of the c-type and the lenticytes are smaller. The chylus cells are found in XVI–XVII (not in XIII–XVI as in F. longeaurita sp. n.), the sperm funnel is shorter (175–200 µm long) with collar as wide as the funnel body. Furthermore, in F. brachiata the oesophageal appendage is of the a-type with only 1–2 short branches, and in the chaetal bundles occur often 4 chaetae postclitellarly both laterally and ventrally (Rota 1994; Schmelz 2003), while in F. longeaurita only 2 or 1 (very rarely 3) chaetae occur behind the clitellum.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Oligochaeta |
|
Order |
|
|
Family |
|
|
Genus |