
Eupolymnia joaoi, Capa, María & Hutchings, Pat A., 2006
|
publication ID |
https://doi.org/ 10.5281/zenodo.174977 |
|
DOI |
https://doi.org/10.5281/zenodo.5657459 |
|
persistent identifier |
https://treatment.plazi.org/id/F84A87E3-7A5E-BD13-B61C-F99BFC97F8CD |
|
treatment provided by |
Plazi |
|
scientific name |
Eupolymnia joaoi |
| status |
sp. nov. |
Eupolymnia joaoi View in CoL sp. nov.
Figures 1 View FIGURE 1 A–G, 2A–C.
Eupolymnia regnans: López et al. 1997: 66 View in CoL , NOT E. regnans Chamberlin, 1919 View in CoL . Type locality. Panama, Coiba National Park, Uvas Island.
Material examined. Panama, Coiba National Park. Holotype: MNCN 16.01/10520, Uvas Island, 7º49'N 81º46'W, 6 m, in filamentous green algae, 7 Feb. 1997. Paratypes – MNCN 16.01/10521, same sample (2 spec.) — USNM 1093374, same sample (4 spec.) — AM W29704, same sample (1 spec. on a SEM stub) — MNCN 16.01/10522 Canal de Afuera Island, 4º41'50''N 81º38'25''W, 6 m, in Caulerpa sertularioides , 29 Nov. 1997 (2 spec.) — AM W29686, same sample (3 spec.).
Comparative material examined. Eupolymnia crassicornis ( Schmarda, 1861) , AM W9137, Bimini, Bahamas, coll. Andrew, 1953 — E. insulana Chamberlin, 1919 , USNM 19357, type, Galapagos Islands, Ecuador, 550m — E. nebulosa ( Montagu, 1818) , BMNH ZK 1950.6.6.21, off Lyme Bay, English Channel, England, 50°10'N 2°55'W — AM W 200882, Looe, Cornwall, England, 18 Mar.1972 — E. regnans Chamberlin, 1919 , MCZ 2395, syntype, Perico Islands, Panama Expedition USFC Albatross, 13 Mar. 1891, coll. Agassiz.
Description. Holotype incomplete, maximum width 3.8 mm; length 11.4 mm for 38 chaetigers. Specimen whitish except for broad band of pigmented eyespots arranged in several irregular rows on compact prostomium. Prostomium forming tentacular ridge from which buccal tentacles arise, although all lost, scars remain ( Figure 1 View FIGURE 1 B, C). Lips well developed, upper lip larger and projected dorsally, lower lip restricted to oral area. Three pairs of branchiae on segments 2–4, with long main stalk and several short dichotomous branches; first pair largest, subsequent pairs progressively smaller; first pair inserted dorso-laterally, second and third pair inserted almost in longitudinal line, although third pair slightly more displaced laterally ( Figure 1 View FIGURE 1 A). Three pairs of lateral lobes on segments 2–4 with second pair largest, arranged in oblique row, with third pair close to first pair of notopodia ( Figure 1 View FIGURE 1 B). First pair elongated, sub-rectangular, wider than long; second and third pairs rounded, wider than long. Fourteen ventral pads, beginning on segment 3, narrowing posteriorly ( Figure 1 View FIGURE 1 C). Nephridial pores on spherical papillae located between noto- and neuropodia of segments 3–8. Notopodia, narrow and elongate, on segments 4–21 ( Figure 1 View FIGURE 1 B). Notochaetae smooth, narrow-winged capillaries, arranged in two irregular rows, graded in length within fascicle ( Figure 1 View FIGURE 1 D, F). Neuropodia from segment 5 continuing to pygidium. Neuropodia with one row of uncini on segments 5–10, subsequently in two rows to segment 17, arranged alternating, face to face. Uncini of the first 17 chaetigers (those with notochaetae) with two rows of teeth over main fang decreasing in size posteriorly, with dental formula MF:2–3:2–3 ( Figure 1 View FIGURE 1 E); dorsal button below main fang acute, situated mid way between prow and base of main fang ( Figure 2 View FIGURE 2 A, B). Neuropodia on segments 18 to pygidium elongate, diminishing in length towards posterior end, with avicular uncini arranged in single rows and dental formula similar to the anterior uncini ( Figures 1 View FIGURE 1 G, 2C). Pygidium not seen.
Var ia t io n. All the paratypes are posteriorly incomplete so details of pygidium are not described. Paratypes are similar in size and colour to the holotype. Prostomial eyespots are conspicuous in all specimens. The branchiae are variably developed in some paratypes and branchial branches may vary in number and length among the specimens but the arrangement and length of the branchial stalk is similar among material studied. In some specimens, nephridial papillae are not visible on anterior segments. Only one specimen (MNCN 16.01/10522) has some tentacles still attached to the prostomium and they are uniformly tapered.
Remarks. There are 20 species of Eupolymnia currently recognised. In some cases, the original descriptions are brief and details about certain features are not provided, and for this reason we have summarised the available data in Table 1 View TABLE 1 , including some diagnostic characters for each nominal taxon. The information has been obtained from literature (original descriptions, redescriptions of type material or descriptions of specimens from type localities) and when possible from material which we have studied (types or specimens from type localities) as indicated in Table 1 View TABLE 1 . Characters such as eyespots or branchial length and ramification are not definitive of species because eyespots can easily fade on preserved material. Branchial morphology can also depend on regeneration processes, environmental conditions or fixation methods. In Eupolymnia , branchiae are ramified, and generally dichotomous, but a variety of terms such as arborescent, dendritic, cauliflower-shape have been used in the literature. We believe that the relative point of insertion of the branchiae is a more useful character and this feature has been included in table 1 when this information was available. The shape and position of the lateral lobes is also a useful character to separate Eupolymnia species ( Hutchings & Glasby 1988), but this character has often been omitted from descriptions ( Table 1 View TABLE 1 ). The dental formula of the anterior and posterior uncini may also be a useful diagnostic character ( Chamberlin 1919b), but in most cases authors have not included these details.
Species of Eupolymnia previously recorded from the Pacific coast of Panama include E. nebulosa from Galeta Reef, Coiba and Taboga Islands ( Monro 1933, Fauchald 1977), and E. regnans from Perico Island and Balboa ( Chamberlin 1919a, Monro 1933, Fauchald 1977). Eupolymnia nebulosa was described originally from Devon, UK, and material from the type locality (AM W 200882) was examined and compared with E. joaoi sp. nov. The two species can be distinguished by the number of ventral pads on anterior chaetigers, 10 in E. nebulosa and 14 in E. joaoi sp. nov., and by the shape of the lateral lobes as E. nebulosa has small and spherical lateral lobes on segments 2 and 4 while the lateral lobe on segment 3 is bilobed and E. joaoi sp. nov. has rounded lobes. A study undertaken by Londoño-Mesa and Carrera-Parra (2005), also records E. nebulosa from the Mexican Caribbean, but further investigations suggest that these specimens could belong to another species (Londoño-Mesa pers. comm.).
Type material of E. regnans was examined (described below) and found to have uncini with one tooth above the main fang ( Figure 2 View FIGURE 2 D, E) while E. joaoi sp. nov., has two rows of teeth above the main fang, both with 2–3 teeth. Eupolymnia joaoi sp. nov. also differs from E. regnans by having 14 ventral pads instead of 18 ( Table 1 View TABLE 1 ). Eupolymnia joaoi sp. nov. has branchiae arranged in a slight oblique line, with first pair dorsalmost, while E. regnans has the three pairs arranged in a longitudinal line. In addition, the lateral lobes are small and pointed in E. regnans whereas they are rounded and larger in E. joaoi sp. nov.
Other species that have been recorded from the Tropical Eastern Pacific are E. insulana described from Galapagos Islands by Chamberlin (1919a) and E. crescentis Chamberlin, 1919 described from California and recorded in Mexico by Rioja (1941) and Bastida-Zabala (1995).
E. regnans MF:1:1 MF:1 middle Perico Island, Chamberlin 1919a MCZ 2395 Panama,? m (holotype)
E. robusta MF:2:1? middle Aru Island, Caullery1944 ZMA V Pol.1561
Indonesia, 13 m (type) E. joaoi sp. nov. MF:2–3:2–3 MF:2–3:2–3 middle Coiba NP, Panama, This paper MNCN 16.01/
0–7 m 10520 (holotype); MNCN 16.01/ 10521, USNM 1093374, AM W29704, MNCN 16.01/10522, AM W29686 (paratypes)
The type locality given is as available in original descriptions. Abbreviations and symbols: loc.
Locality; lat. Lateral; nephr. Nephridial; pap. Papillae, n. number; ant. anterior; post. posterior; unc.
Uncini;? no information found; + present; - absent; # no type material exists; x (dental formula):
row with one or more teeth, information obtained from lateral view drawing.
The original description of E. insulana ( Chamberlin 1919a) is incomplete and with only the chaetae illustrated, and the species appears not to have been recorded subsequently. Type material of this species was examined (USNM 19357) and, although it was in poor condition and no branchiae remain, it can be distinguished from E. joaoi sp. nov., by the dental formula of the anterior uncini, with only one row of teeth above the main fang in E. insulana , and two rows above the main fang in E. joaoi sp. nov. The depth at which it was recorded (550 m) makes it unlikely that it is conspecific with E. joaoi sp. nov., which was collected in shallow water.
Similarly, E. joaoi sp. nov., can be distinguished from E. crescentis by the dental formula of uncini as both anterior and posterior uncini have only one row of teeth over the main fang in the latter species ( Chamberlin 1919 b), whereas E. joaoi sp. nov., uncini always have two rows of teeth over the main fang.
In the Grand Caribbean Region, another species of Eupolymnia , E. crassicornis , is common in shallow water. These species can be distinguished by the presence of nephridial papillae on segments 3–8 in E. joaoi versus 3–5 in E. crassicornis , and also by the presence of uncini arranged in single rows on segments 5–10 in E. joaoi while they occur on segments 5–9 in E. crassicornis . These two species can also be separated by the dental formulae of the uncini.
Etymology. This species name is dedicated to João Miguel de Matos Nogueira, for his valuable taxonomic research on polychaetes.
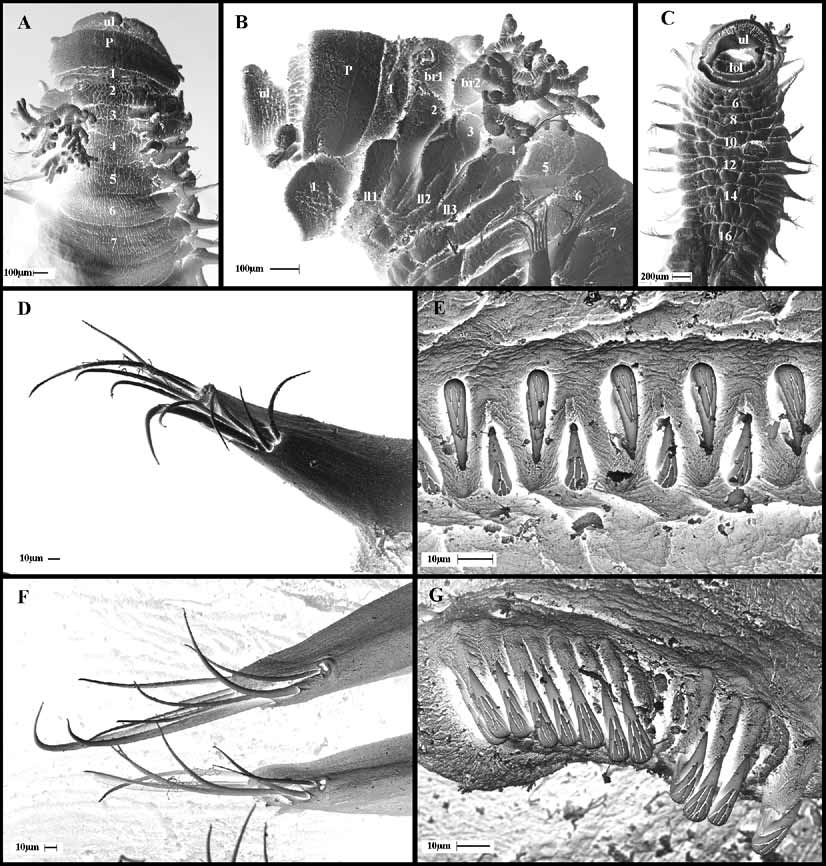

TABLE 1. Eupolymnia species diagnostic characters.
| Taxon | Eye- spots | Lat. lobe shape/size | Lat. lobe arrangement | Branchial shape/ size | Branchial arrangement | Neph. pap. (segs.) | Ventral pads (n.) | Unc. single rows (segs.) |
|---|---|---|---|---|---|---|---|---|
| E. boniniana (Hessle, 1917) | + | small, 1st largest | oblique | 1st longest | longitudinal | 2–4 | 14 or 15 | ? |
| E. capensis (McIntosh, 1925) | + | ? | ? | 1st longest | ? | ? | ? | ? |
| E. congruens (Marenzeller, 1884) | + | 3rd with prolonged margin | 1st ventral | short thick stalks, 1st longest | ? | ? | 16 | 5–10 |
| E. crassicornis (Schmarda, 1861) | + | 1st largest | oblique, 3rd dorsalmost | long stalks; 1st largest | oblique, 3rd dorsalmost | 3–5 | 14 or 15 | 5–9 |
| E. crescentis Chamberlin, 1919 b | + | ? | ? | long and thick stalks, 1st longest | ? | 3–5 | 14 or 15 | 5–10 |
| E. dubia (Caullery, 1944) | + | small | oblique, 3rd closest to notopodia | 1st longest, long stalk | longitudinal | ? | 14 | 5–9 |
| E. heterobranchia (Johnson, 1901) | - | 1st largest, connected ventrally, rest small | oblique, 3rd dorsalmost | 1st longest, long stalk | 2nd dorsalmost | 3–5 | 14 | 5–11 |
| E. insulana Chamberlin, 1919 | - | present | 2nd dorsalmost | ? | ? | 3–8 | ? | 5–10 |
| E. intoshi (Caullery, 1944) | + | 2nd largest | oblique | 1st longest, short stalk, | 3rd dorsalmost | 2–3 | ?14 | 5–10 |
| E. kermadecensis (McIntosh, 1885) | ? | ? | ? | only 1st pair present, with long stalk | ? | ? | ? | ? |
| E. koorangia Hutchings & Glasby, 1988 | + | rounded, 1st connected ventrally | oblique, 3rd dorsalmost | thick and short stalk | 2nd dorsalmost | 3–5 | 14 | 5–10 |
| E. labiata (Willey, 1905) | ? | ?3rd largest | ? | ? | ? | ? | ? | ? |
| E. marenzelleri (Caullery, 1944) | + | crenulated margins, 2nd largest | 2nd dorsalmost | short stalk, small, decreasing posteriorly | ? | 5 | 15 | ? |
| E. nebulosa (Montagu, 1818) | + | 2nd largest and bilobed | oblique | no stalk, few branches | ? | 3–9 | 10 | 5–10 |
| E. nesidensis (delle Chiaje, 1828) | + | ? | ? | ? | ? | ? | ? | ? |
| E. regnans Chamberlin, 1919 | + | pointed, small | oblique, 3rd dorsalmost | long stalk, 1st longest | longitudinal | 3–8 | 18 | 5–10 |
| E.robusta (Caullery, 1944) | + | rounded, large | oblique, 3rd dorsalmost | long stalk, 1st longest | oblique, 1st dorsalmost | 3–5 | ? | 5–9 |
| E. trigonostoma (Schmarda, 1861) | - | ? | ? | long | ? | ? | ? | ? |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Eupolymnia joaoi
| Capa, María & Hutchings, Pat A. 2006 |
Eupolymnia regnans: López et al. 1997 : 66
| Lopez 1997: 66 |