
Trichopterygini Warren, 1894
|
publication ID |
https://doi.org/ 10.5281/zenodo.279481 |
|
DOI |
https://doi.org/10.5281/zenodo.6184077 |
|
persistent identifier |
https://treatment.plazi.org/id/D2566362-FFAF-FFF9-FF75-FF0BDECEFBBB |
|
treatment provided by |
Plazi |
|
scientific name |
Trichopterygini Warren, 1894 |
| status |
|
Tribe Trichopterygini Warren, 1894
( Figs 22–30 View FIGURES 22 – 30 )
Pierce (1914) was the first to present a definition of the group (as Lobophorinae); unfortunately he also grouped two genera of the tribe Triphosini with. The genera combined now in Trichopterygini appear to be characterized by a wide ostium, and corpus bursae ornamented with petaloid signa. The variation of wing venation and sexual differences within species are markedly large.
The tribe is relatively well studied. Dugdale (1980) described its Australian taxa and explained the modification of male sternite A2 as a shelter for androconial pencils. Parra (e.g. 1996) studied the diversity of trichopterygine genera in Chile and Holloway (1997) that in the Indo-Australian region. Holloway (1997: 100) characterizes Trichopterygini by the reduction of the anal area of male hind wing to fold, crevice, flap or lobe(s); the 2nd sternite modified in male, having an internal, hollow sack; valva in male genitalia has both costal and saccular ornamentation; bursa copulatrix is generally scobinate (shared with Chesiadini , Eupitheciini , Melanthiini etc.), the character also occurs in Idaeini ( Sterrhinae ).
Trichopterygini shares the trend of modification of hind wing venation with Dyspteridini and Chesiadini . The tribe is a composite of many deeply and differently specialized elements, maintaining also archaic characters like the presence of gnathos or gnathi in some East Asiatic genera ( Xue & Zhu 1999), or the occurrence of lateral appendages to juxta in a Chilean genus. Being speciose in Neotropical and Oriental faunas, only two lineages (one with the uncus conical and valva apex produced a spine, another with the uncus distal projection long and valva indented distally) have Holarctic distribution.
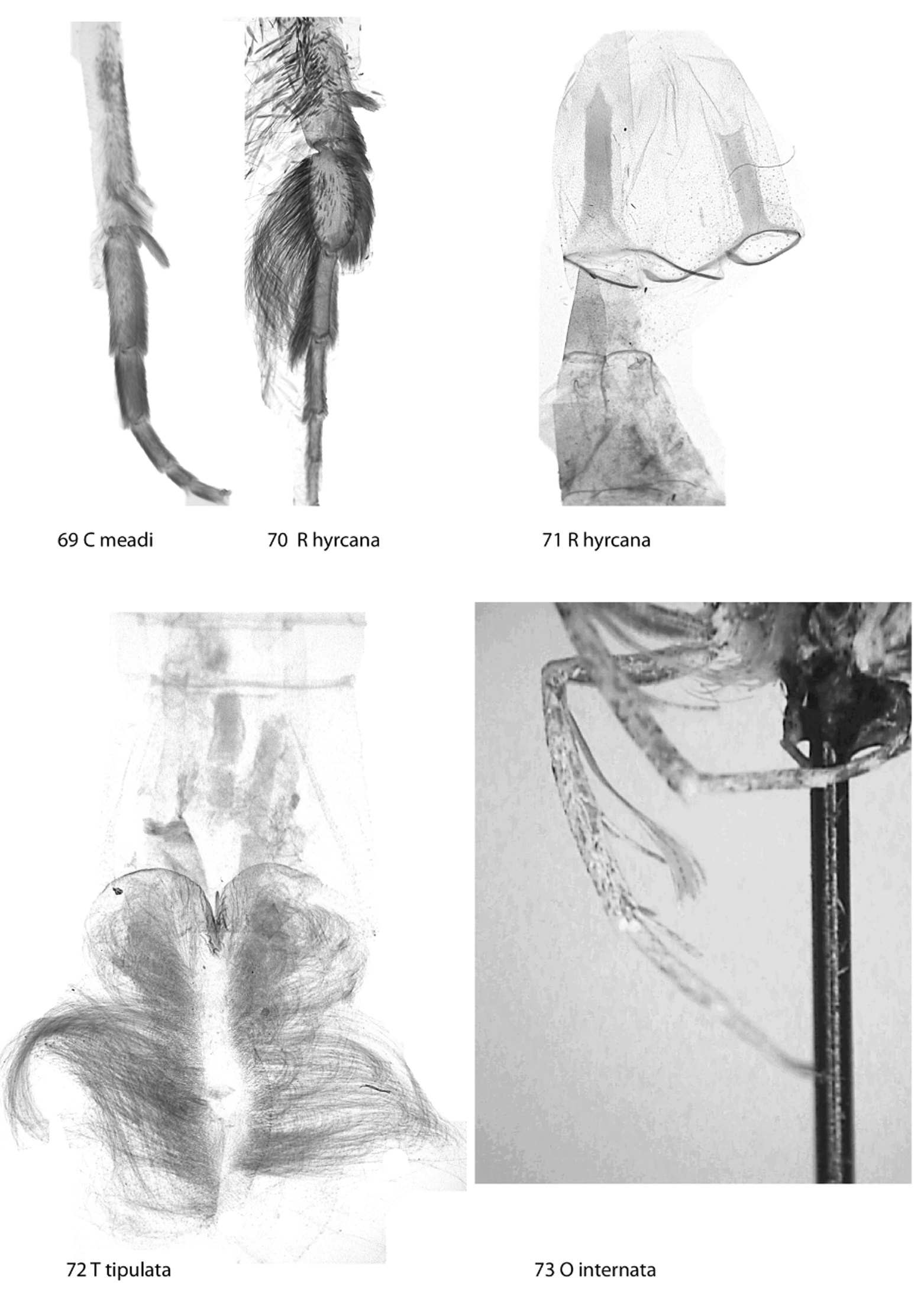
Dugdale (1980) and Xue et al. (2008) pay attention to the modification of male hind leg characteristic of this tribe: sometimes there is a thin tibial hair pencil ( Fig. 73 View FIGURES 69 – 73 ), and the trend of spur reduction has genus-level significance. However, there are at least three trends of modification, partly overlapping. Two genera, Tatosoma Butler , and Tricalcaria Han , have lost one of the medial spurs on the normally long hind legs. The Sauris Guenée group of genera has proximal spurs or all spurs on male hind tibia reduced and various hair tufts present (see Dugdale 1980). A thin and long hair pencil in some Holarctic genera, such as Lobophora Curtis , Oulobophora Staudinger a.o., attached to the base of male hind tibia and concealed in sternite A2 makes out the third trait ( Fig. 73 View FIGURES 69 – 73 ). The tribe is diverse and some interesting characters occur within subclades: hind wing costal venation often different between sexes; subapical thickening or clubbing of male antennae which are apically sickle-shaped; very long abdomen in male; modifications of vestiture and scaling of wings and abdomen ( Fig. 72 View FIGURES 69 – 73 ); occurrence of abdominal or valval coremata, usually concealed in the hollow structure of sternite A2, but also as hair pencils or scale tufts on male hind legs.
Distribution: Neotropical, Indo-Australian, Afrotropical, Holarctic. The morphological diversity of genera seems to be the greatest in South America (e.g. Parra 1996) and East Asia ( Yamamoto and Sota 2007; Choi 2008).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |