
Eulimnogammarus cyanoides ( Sowinsky, 1915 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3838.5.2 |
|
publication LSID |
lsid:zoobank.org:pub:33D621CE-0856-42F6-BEA9-A75A458ABCE4 |
|
DOI |
https://doi.org/10.5281/zenodo.6140225 |
|
persistent identifier |
https://treatment.plazi.org/id/BC2587EA-E07C-5D60-2BFF-BD649D5B949D |
|
treatment provided by |
Plazi |
|
scientific name |
Eulimnogammarus cyanoides ( Sowinsky, 1915 ) |
| status |
|
Eulimnogammarus cyanoides ( Sowinsky, 1915) View in CoL
Figures 2–7 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7
Echinogammarus cyanoides . Sowinsky 1915, 154–155, figs 155, 156; Taf. XX, figs 16–19.
Non Eulimnogammarus cyanoides . Bazikalova 1945, 209–210; Taf. XXV, fig. 3; Taf. XXVI, fig. 1.
Material examined. Lectotype: female, 9.3 mm, ZIN 1/68949; Russia, Lake Baikal, 12.08.1902, collected near the Solzan River at a depth of 1−3 sazhen (1 sazhen = 2.16 m). The lectotype is designated here.
Paralectotypes: female 8.7 mm, ZIN 2/88512, from the same sample as the lectotype; male 10.2 mm and 1 dissected specimen, ZMK No. 356 b, the same sample as the lectotype.
In addition, we determined that two further dissected specimens from the original sampling of Sowinsky did not belong to this species: 1 sp., ZMK No. 356 b, from the same sampling; 1 sp., ZMK No. 356 c, 15.06.1900, Lake Baikal, collected by a diver in Maly Baranchik Bay at a the depth of 3−4 sazhen.
All specimens in ZMK were in the status of syntypes ( Kostyuk 1973); they are also marked as syntypes in a museum digital catalogue (http: // museumkiev.org / zoology / catalog / cat_ types _titul.html). Earlier (not later than 1989), an unknown person selected a lectotype and paralectotypes from this series of syntypes. Nevertheless, the lectotype status had not previously been published and therefore is not considered valid. The specimen, marked by the unknown person as a lectotype (collected from Baranchik Bay, ZMK No 356 c), does not belong to the species E. cyanoides , because it has the following characteristics: long setae already present on the second pleon segment whereas E. cyanoides has long setae that begin from the third segment. The eye is strongly curved, the lower part of which is markedly wider than the higher part, and its vertical diameter is 2.5-fold bigger than horizontal diameter. The basal article of the peduncle in antenna 1 widens in the middle. The propodites of the gnatopod 1 are small (even in female of E. cyanoides they are very large) and shorter than the head and even shorter than the propodites of the gnatopod 2. The taxonomic status of this specimen must be defined separately; however, it cannot be considered a lectotype for E. cyanoides . One specimen from the collection unit ZMK No. 356 b also shows setae beginning from the second segment of the pleon in addition to two groups of spines in the third segment of the pleon, which strongly indicates that this specimen belongs to another species.
Due to the ICZN requirements, the lectotype is designated in this report. Sowinsky (1915) indicated “Solzan” as the first site where the species was noted; we therefore assigned the lectotype from a number of syntypes collected at this place.
There is one E. cyanoides specimen in the “Catalogue of amphipod collection (including type specimens) stored in the Limnological institute SB RAS” ( Mekhanikova et al. 2010 –2011), which was identified by Dorogostaisky and was collected from the Ushkanji islands region (№ 670, 12.08.1921). The revision of this one specimen revealed that it does not belong to the species E. cyanoides because segment 3 of the pleon is armed with 3 lines of spines, which contradicts the diagnosis. Its eyes are also quite wide.
Diagnosis. Four posterior body segments with very long setae. Only segments of urosome armed with spines. Eyes wide-reniform, their height not more than twice time more then width. Antenna 1 exceeds 2/3 of body length; basal article of the peduncle distally with long thin posteromedial spine or two spines of varying lengths. Peduncles of antennae 1 and 2 with sparse bunches of setae. Gnathopod 1 propodi large, 1.5 times exceeding head length. Posterior-inferior angles of bases 6 and 7 without lobes. Uropods 1 and 2 with sparse long spines. Uropod 3 with sparse simple setae; outer ramus with rudimentary second article. Telson lobes with spines only on their apexes. Body length of adult females approximately 9 mm.
Description. Female. Body smooth; habitus Gammarus -like (compact, laterally narrowed). Pleonites 1 and 2 without setae or spines. The 3rd segment of pleon with many long setae in geminate-groups on the dorsal surface and along posterior margin; the longest setae markedly greater in length than urosomal segment 1. All three urosomites bear twin groups of 1−3 long, thin spines that are suberected or almost completely erected, and long setae (greater than length of segment bearing them) on posterior margin.
Head slightly swollen from dorsally and terminated by very short rostrum; interantennal lobe poorly developed and smoothly rounded. Eyes dark and wide-reniform; their vertical diameter exceeding horizontal diameter by approximately 2-fold and is slightly larger than diameter of basal article of antenna 1 peduncle.
Antenna 1 is slightly longer than 2/3 of body length. Basal article approximately the length of head. On its anterior-inferior angle, it bears one long, thin spine or two spines—a long one and one that is 0.5−0.75 times as long as the other (cf. Fig. 3 View FIGURE 3 , 7 View FIGURE 7 ). Inferior margin with a single seta (lectotype) or in one case, a single small thorn was observed ( Fig. 7 View FIGURE 7 ). Article 2 is marginally longer than basal article; article 3 is <0.7 times as long as article 2; both have sparse setae on their inferior margin, longer in distal portion. Primary flagellum 20−30-articulate. Accessory flagellum with 3−4 fully developed and one rudimentary articles.
Basal article of antenna 2 swollen, bearing 0−2 setae; antennal cone thin and sharp and slightly shorter than article 3 of peduncle; articles 4 and 5 long and thin, whereas article 5 is 0.75 times as long as article 4, and both bearing bunches of sparse but long setae on their inferior margin. Flagella with 9−10 articles noted bearing sparse, long setae. Calceoli absent.
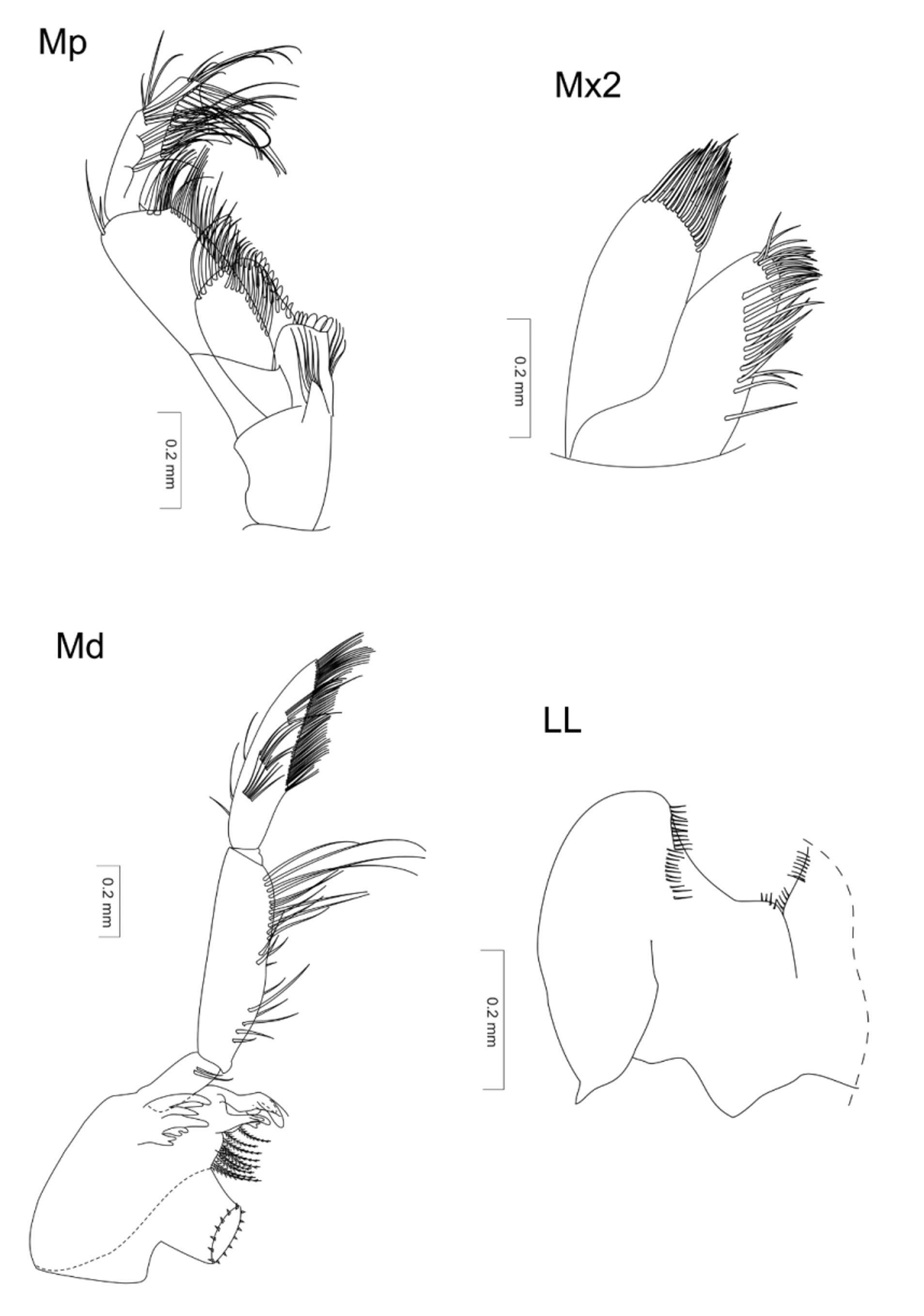
Mandibles. Incisors with 6 denticles, and dental row comprising 4 setae. Article 1 in the mandibular palp with 2 setae (one very short); article 2 wide with long setae on distal third; article 3 lanceolate with a brush (D3-setae) that is 2/3 its length with equal-length setae.
Maxilla 1 (In lectotype damaged). Outer plate with 8 spine-teeth, two of which bifurcated, and others obliquepectinate. Inner plate bearing 11−16 plumose setae. Palp narrower than outer plate, extending over apexes of spineteeth on this plate. Distal end of palp with 2 setae.
Maxilla 2 plates elongated, of equal size, and inner plate bearing 14 plumose setae on oblique row.
Maxilliped. Outer plate wide, exceeding middle of third palp article; inner plate small, not exceeding middle of outer plate length. It bears 3 blunt, tapered teeth. Terminal article of palp slightly shorter than middle article and amplified on its apical part; dactylus thin and elongated, in proximal portion half as wide as terminal palp article (its distal portion).
Lower lip: lobes wide, short; setules of inner margins poorly developed.
Coxal plates bare; coxa 1 wide, tetragonal and shorter than coxa 2, with skewed anterior and posterior margins. Coxae 2 and 3 linguiform; coxa 4 with smooth, rounded lobes in the inferior 1/3 of posterior margins. Propodus of gnathopod 1 very large, oval, 1.5 times as long as head and twice as long as propodus of gnathopod 2. Palmar margin markedly oblique, and inferior margin swollen, with bunches of sparse short setae. Proximal 1/3 of palmar margin with thick, blunt spine together with group of boundary spines. Inferior margin without spines. Dactylus very curved. Carpus of gnathopod 1 short.
Propodus of gnathopod 2 weak, tetragonal, and its anterior and posterior margins parallel. Palm margin short and swollen. Only boundary spines present. Posterior margin without spines, bearing dense setae. Carpus elongated.
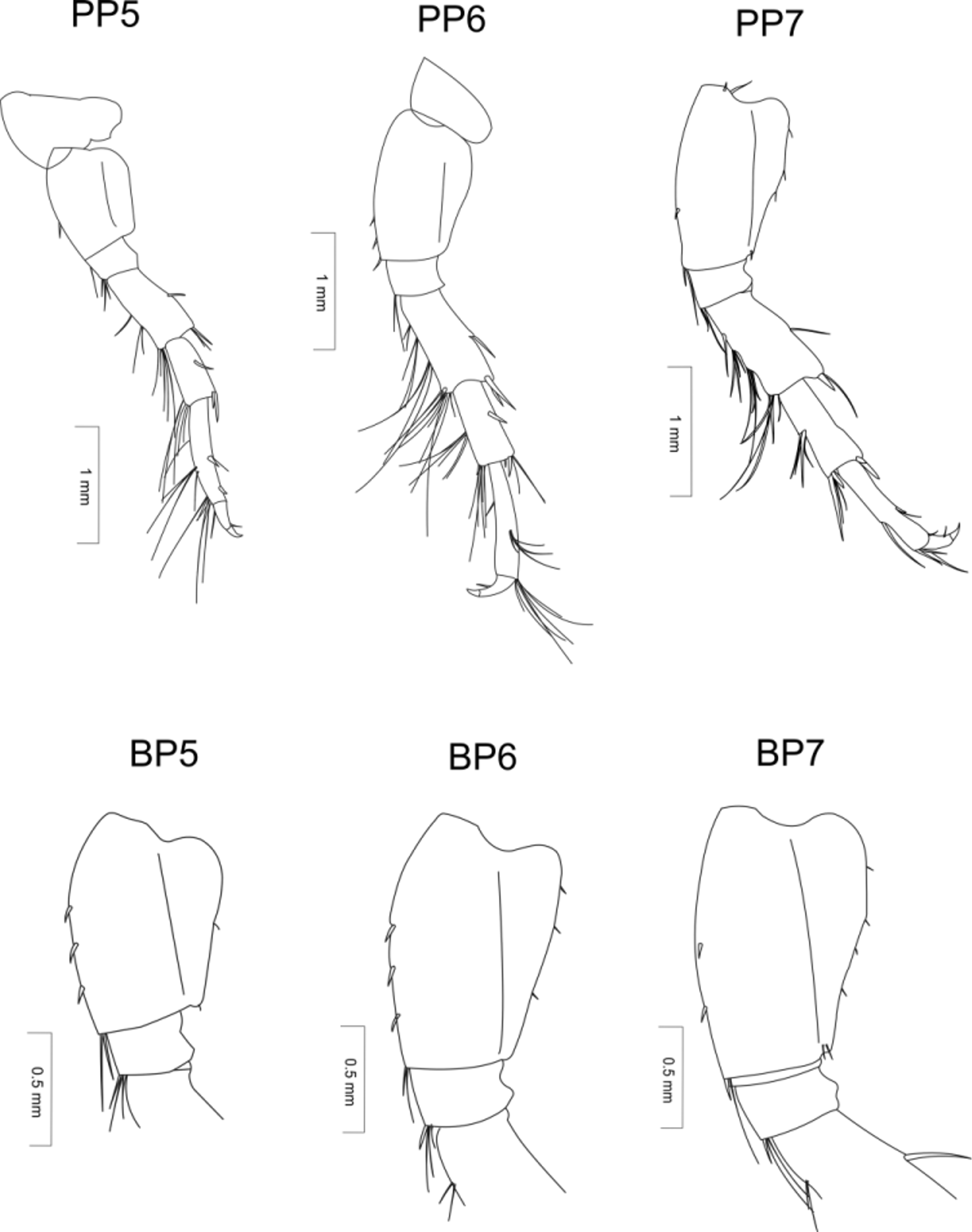
Pereopods 3 and 4 short, with sparse, long setae and only single spines. Meri longer and significantly wider than carpi and are approximately as long as propodi. Dactyli short, thick and curved.
Pereopods 5−7 rather short. Pereopods 5 noticeably shorter than pereopods 6. Pereopods 7 slightly shorter than pereopods 6, articles bearing sparse, long spines and sparse bunches of setae, particularly long on anterior margins of articles. Bases of all three pairs bearing a single, short seta on their anterior and posterior margins. Length of basis 5 is 1.5 times width; posterior margin distinct, forming obtuse lobe. Basis of pereopod 6 more elongated, and its posterior margin slightly cut and its inferior portion not developed. Distal parts of merus and carpus of pereopods 5−7 twice as wide as proximal part of propodus. Propodi thin, slightly curved and not less than 1.5 times longer than carpi. Distal ends of propodi in pereopods 5 and 6 with bunches of very long setae; fifth propodi with setae along entire length of propodus. Dactyli length approximately 1/3 the length of propodi, robust and curved at apices.
Epimera of pleonal segments almost unarmed; epimera 1 short and rounded; epimera 2 and 3 wide, without short spines along their inferior margin. Inferior-posterior angle of epimera 2 slightly dulled and in epimera 3 acuminated.
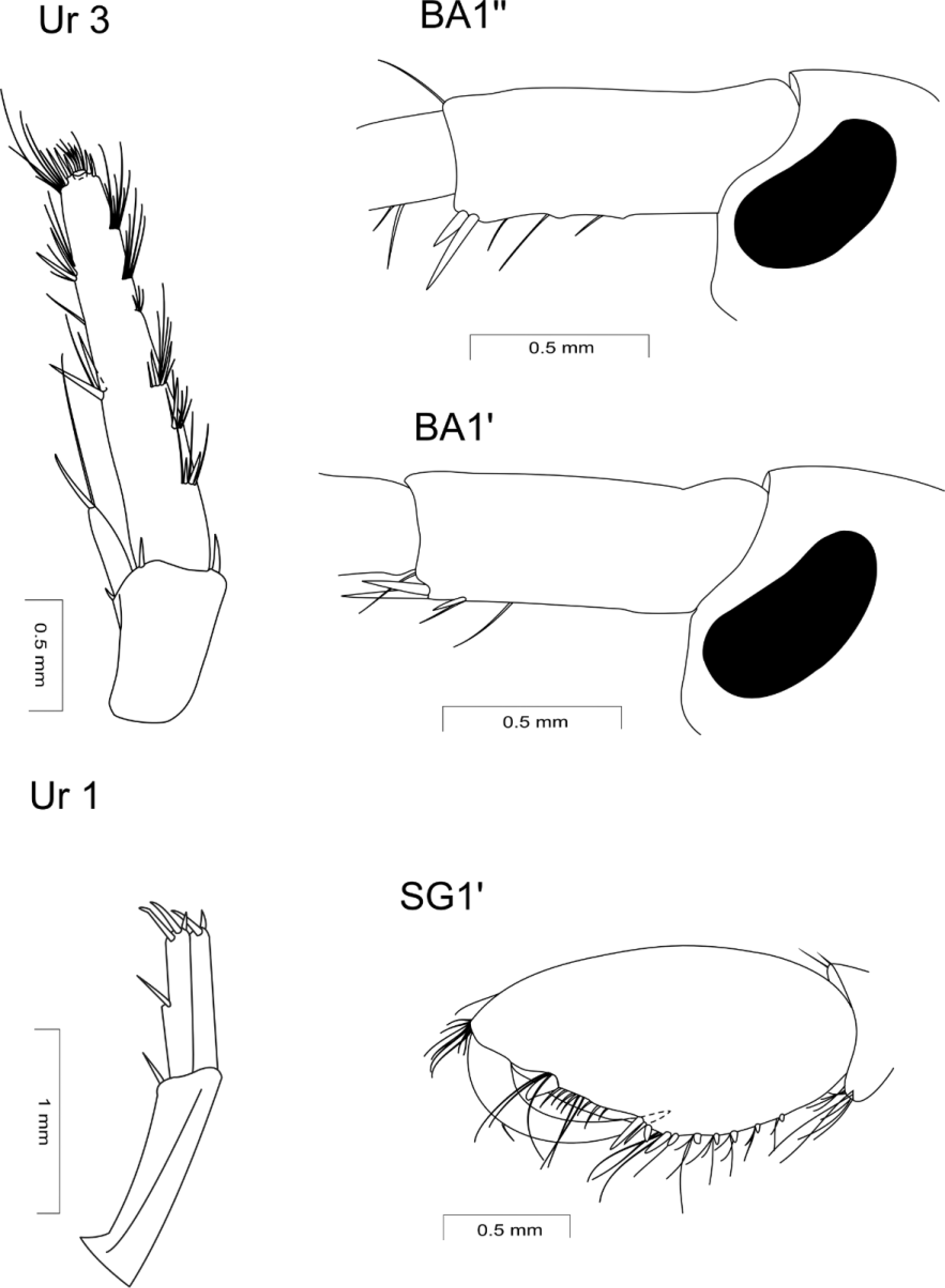
Uropods 1 and 2 protruding backwards and of similar lengths, quite thin and armed by single, long, thin spines. Rami of uropod 1 equal in length; outer ramus of uropod 2 is 0.75 times as long as inner ramus; inner rami only with apical spines, whereas outer rami bearing additional medial spines.
Uropod 3 is 1/4 the length of body; outer ramus 3 times as long as peduncle and 4−4.5 as long as inner ramus. It bears a rudimentary second article and sparse long spines and bunches of simple setae on both margins and apex (parts of setae apparently broken). Inner ramus with one long spine and simple seta on apex.
Telson cleft up to the base; telson lobes tapering, with 1−2 spines and 2−3 setae; lateral margins of lobes with 1 seta each.
Male: Inferior margin of propodus of gnathopod 1 with a row of short and rather wide, tapering thorns beginning from 1/3 of article’s length ( Fig. 7 View FIGURE 7 ). Propodus of gnathopod 2 narrower than in female.
Body length 8.0− 10.2 mm (our measurements). Sowinsky (1915) described the body length as being 12−13 mm. Specimens of this latter length are absent among the type specimens, recognized by us as belonging to this species.
Comparative remarks. Based on a number of characteristics, this species corresponds to the later-described E. grandimanus Bazikalova, 1945 . First, presence of spines only on three urosomital segments; the low eyes; presence of a spine on the peduncular article of antenna 1; and the characteristic large propodi of gnathopod 1 (see: Bazikalova 1945, taf. XXVII, fig. 2). It is notable that the propodus of gnathopod 1, depicted in this figure by Bazikalova, bears a hard, blunt thorn in the middle of the palmar margin and a group of the boundary thorns, which, as indicated above, is characteristic for female of E. cyanoides . And more, according to this description the lower margin carries paired blunt, short, small thorns, characteristic for male specimens of E. cyanoides (page 212).
Using the key of Bazikalova (1945), all of the examined type specimens of E. cyanoides were identified as E. grandimanus .
However, Bazikalova (1945) indicated great variability for E. grandimanus , which shows variation in the arming of the third pleonal segment (setae are absent in some cases but are dense, long and form two rows in other cases). Variability was also described for the uropod 3 structure (length of rami and vastly different densities of setae). These differences indicate that E. grandimanus is initially presumed to be a composite species. We assume that E. grandimanus sensu Bazikalova is a minor synonym of E. cyanoides . The revision of its holotype at the ZIN (1/50177) in 2013 showed that uropod 3 of the E. grandimanus holotype is equipped with curved simple setae, whereas no one of E. cyanoides are curved. A detailed study of the intraspecific polymorphisms of E. grandimanus sensu Bazikalova is necessary for a final decision to synonymize E. grandimanus with E. cyanoides or to divide first species into two or more species.
Examination of the available specimens of E. grandimanus , collected in different areas of Lake Baikal (Bolshie Koty Bay, Cape Bolshoy Kadilnyi, Cape Pokoyniki, Bolshoy Ushkaniy Island) revealed the following inconsistent differences from the type specimens of E. cyanoides : setae on the last four body segments of E. grandimanus do not lay backwards, but are rather upright and often bend forward; basal article of the antenna 1 peduncle, articles of antenna 2 and its flagellum are comparatively shorter and thicker. Even in large (8−10 mm) specimens, the secondary flagellum comprises 2−3 distinct articles, one of which is rudimentary (if any exists). Pereopods 3 and 4 bear bunches of very short setae. The rami of uropods 1 and 2 are shorter. The outer ramus of uropod 3 is also significantly shorter than in E. cyanoides , and the setae vary in length and density; however, in most cases, they are more or less curved (as in the holotype).
The differences comparison with other morphologically close species— E. cyaneus (Dybowsky) and E. cyanellus Bazikalova—that could be confused with E. cyanoides are provided below. In all three species, only the last three urosomites are armed with spines and setae.
E. cyanoides is different from E. cyaneus based on the following characteristics: 1. Setae on the third pleonal and on the first and second urosomal segments are significantly longer. 2. Setae on both antennae are less dense and are collected in a smaller number of bunches. The inferior-anterior angle of antenna 1 bears a long thorn without dense setae. 3. The terminal article of the mandibular palp carries a brush with setae of equal length (the setae in a brush of E. cyaneus are unequal). 4. All pereopods show less setae, and the posterior margin of the basis in pereopod 7 is poorly developed.
E. cyanoides differs from E. cyanellus based on the following characteristics: 1. The inferior part of the eyes does not extend (in E. cyanellus , they are extended). 2. The setae on the last 4 body segments are quite dense and long (in E. cyanellus , they are sparse and short). 3. The basis of pereopod 5 is slightly concave on the posterior margin (in E. cyanellus , this margin is convex).
However, besides spine armor being only on the final three segments of the body, both species show other characteristics that are similar: the structure of the basal article of antenna 1 (inferior-anterior angle bears 1−2 spines), uropods 1 and 2 (rami of the first pair are equal, the outer ramus of the second pair is shorter than the inner ramus) and the basis of pereopod 7—in E. cyanellus , the inferior-posterior angle protrudes slightly whereas in E. cyanoides , it is smoothly rounded (see Fig. 6 View FIGURE 6 ).
Distribution and ecology. Russia, Lake Baikal. Sowinsky (1915) described the following sampling points. 1. The Solzan River at a depth of 1−3 sazhen (approximately 2−7 m), stones, sand. 2. Maly Baranchik Bay, collected from sponges (at a depth of 3−4 sazhen). 3. Ushkanyi islands. Only the first point can be considered accurate. Identification of this species from the second sampling point is inaccurate (see above). It is necessary to consider that only a limited number of species show biocoenotic relationships with sponges in Lake Baikal. The third sampling point, from which no intact samples remain, may also be erroneous. As described by Sowinsky (1915, p. 155): “Ushkanyi islands. Dredged from a depth of 20 sazhen (43 m), 12 specimens ”. However, a list of all of the species found in the description of the obtained material from the different areas of the lake (p. 60) exists, which states for the area of Ushkanyi islands: “ Echinogammarus cyaneus (Dyb.) —12 ex.”. Nevertheless E. cyanoides is absent from the list. However, the presence of E. cyaneus at that depth remains doubtful because this species inhabits only narrow areas of the water edge and can also be found up to depths of 5 m ( Bazikalova 1945; Weinberg et al. 2002; Govorukhina 2005, etc.).
After the re-examination of E. cyanoides specimens from the area of Solzan, it was found that one of the specimens (lectotype, ZIN 1/68949) was a female of the fifth stage and another (paralectotype, ZIN 2/88512) was a female of the second stage, indicating that at the moment of sampling, this species was approaching the end of the reproduction period.
As stated above, Bazikalova (1945) described, under the name of E. cyanoides , another species that appears to be E. messerschmidtii sp. n. (original materials of Bazikalova are absent in LIN and ZIN). Therefore, it is unlikely that the reports of Bazikalova (1945) and of other authors ( Golyshkina 1963; Kamaltynov 2001) on the new locations of E. cyanoides are accurate. Obviously the determination of this species by Golyshkina (1963) for silts with the detritus in the Irkutsk water reservoir is erroneous. However, if E. cyanoides ( Sowinsky, 1915) and some representatives of E. grandimanus ( Bazikalova, 1945) are synonymous, this species may be considered widespread in the Baikal littoral zone (0.5–15 m).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |