
Diunatans luctoretemergo, Bosselaers & Post, 2010
|
publication ID |
https://doi.org/ 10.5252/g2010n2a6 |
|
persistent identifier |
https://treatment.plazi.org/id/03AC330B-FFD1-DC5F-FF35-D178BA4FFACA |
|
treatment provided by |
Marcus |
|
scientific name |
Diunatans luctoretemergo |
| status |
sp. nov. |
Diunatans luctoretemergo n. sp.
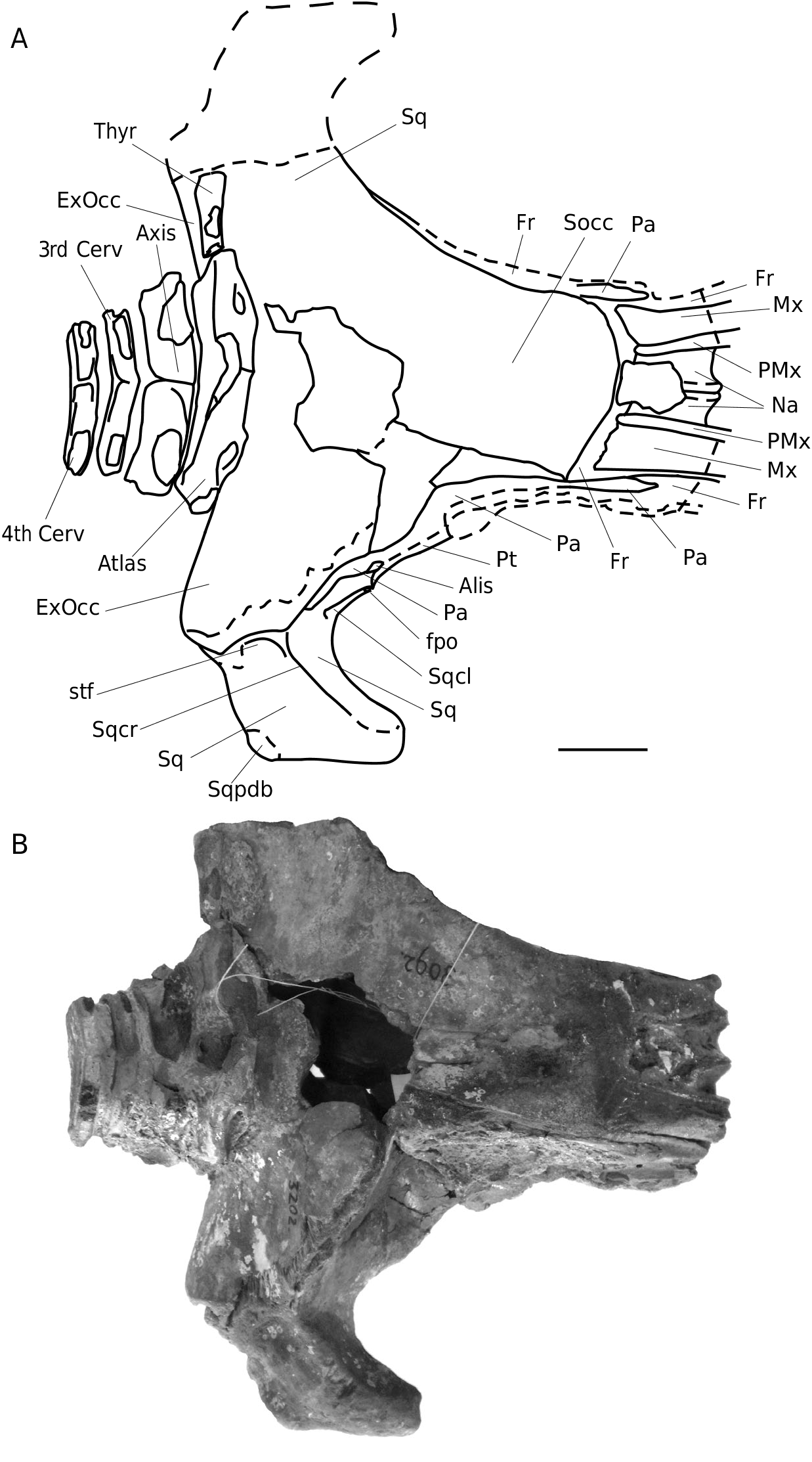
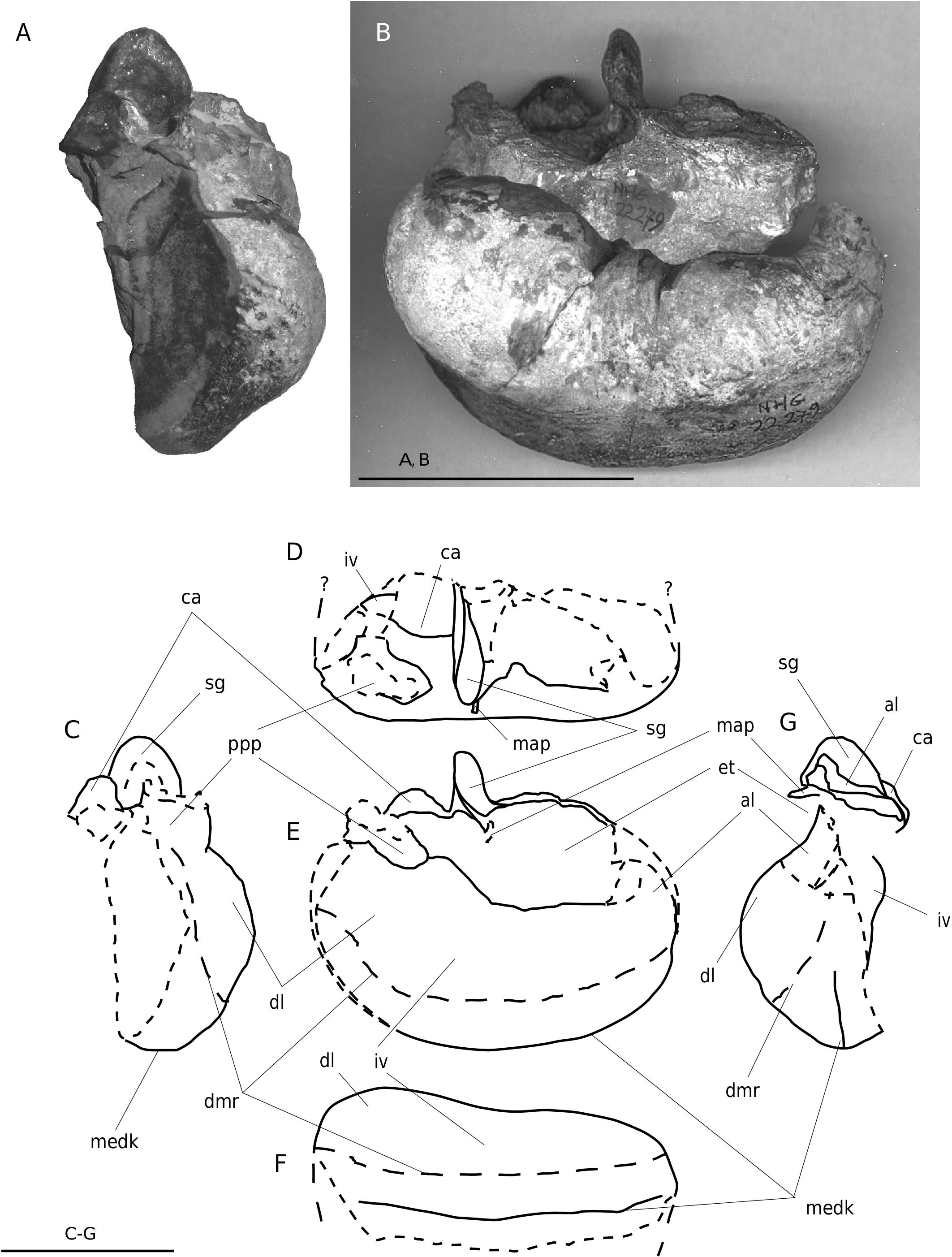
HOLOTYPE. — NHG 22279: neurocranium with nasals, ascending fragments of maxilla and premaxilla, damaged frontals (the lateral part of the sop is missing), parietals, occipitals, squamosals, petrotympanic complexes, fragment of hyoid and four cervical vertebrae. Most of the rostral elements anterior to the vertex, and the dentaries, are missing ( Figs 2-4 View FIG View FIG View FIG ; 7-9 View FIG View FIG View FIG ; 11; 12 View FIG ).
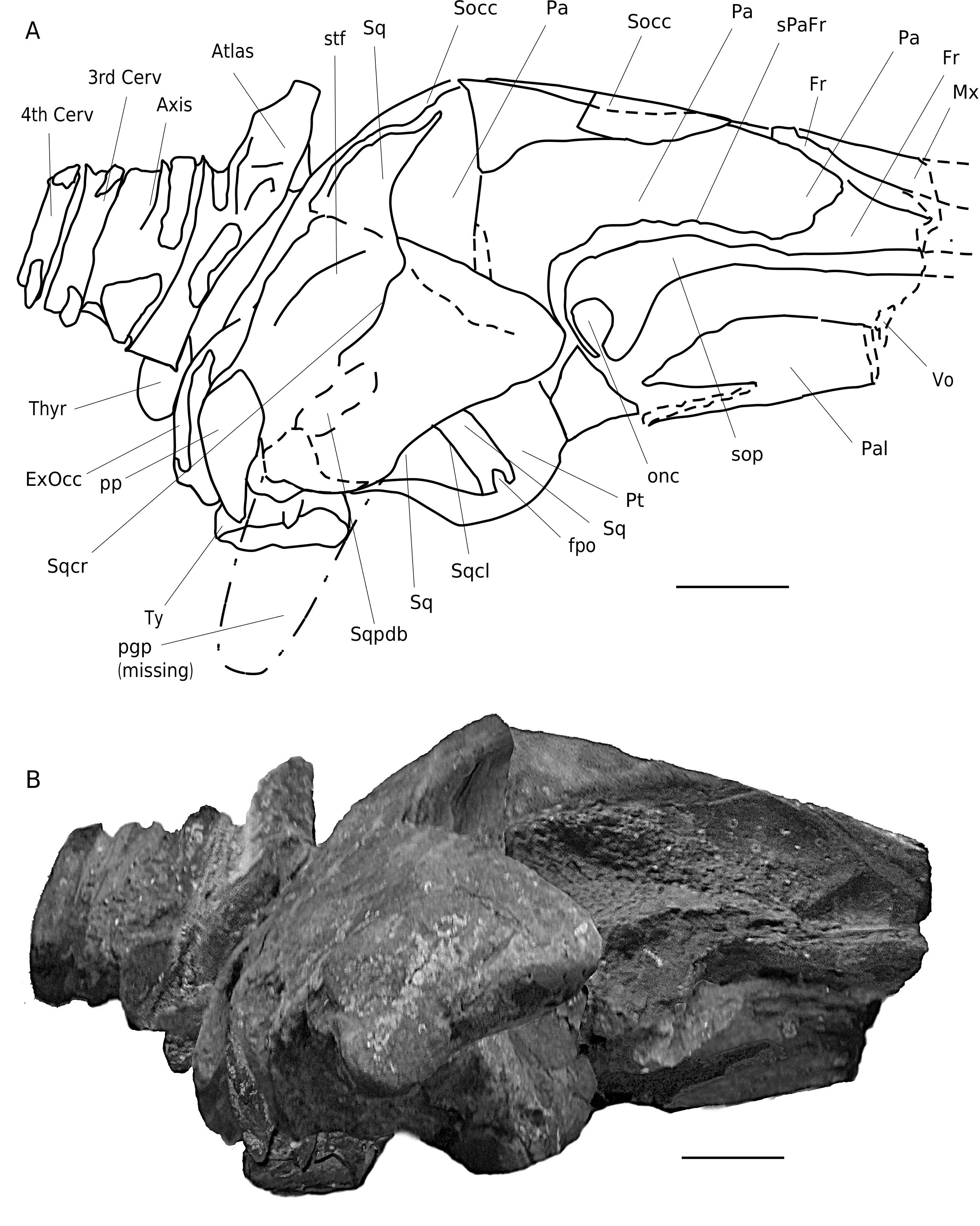
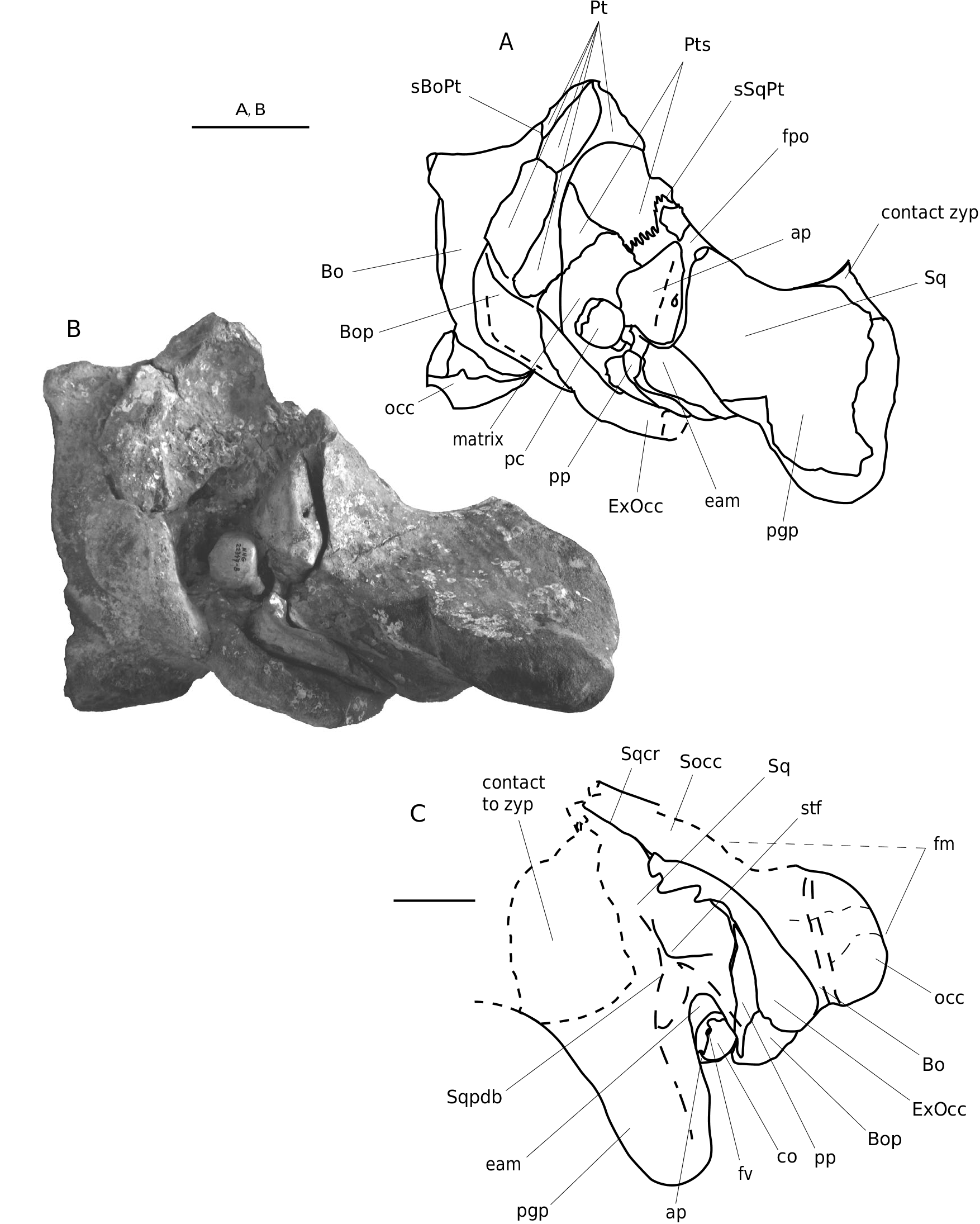
PARATYPE. — NHG 22347: damaged neurocranium with left occipital condyle, left half of basicranium and basisphenoid, left pterygoid, left periotic (the tympanic bulla is missing), left squamosal, left half of supraoccipital, and the posterior process of palatine (overlapping the pterygoid) ( Figs 5; 6 View FIG ; 10 View FIG ) .
ETYMOLOGY. — After the motto “ Luctor et Emergo ” of the Dutch province Zeeland where holotype and paratype specimens were collected and rediscovered, thus symbolising the rise of the specimens above the sea surface and – perhaps even more spectacular – their survival after being forgotten for 70 years on dusty museum shelves.
DIAGNOSIS. — As for genus.
HORIZON AND AGE. — Zanclean, Kattendijk Formation, Kattendijk Sand Member (?), Early Pliocene.
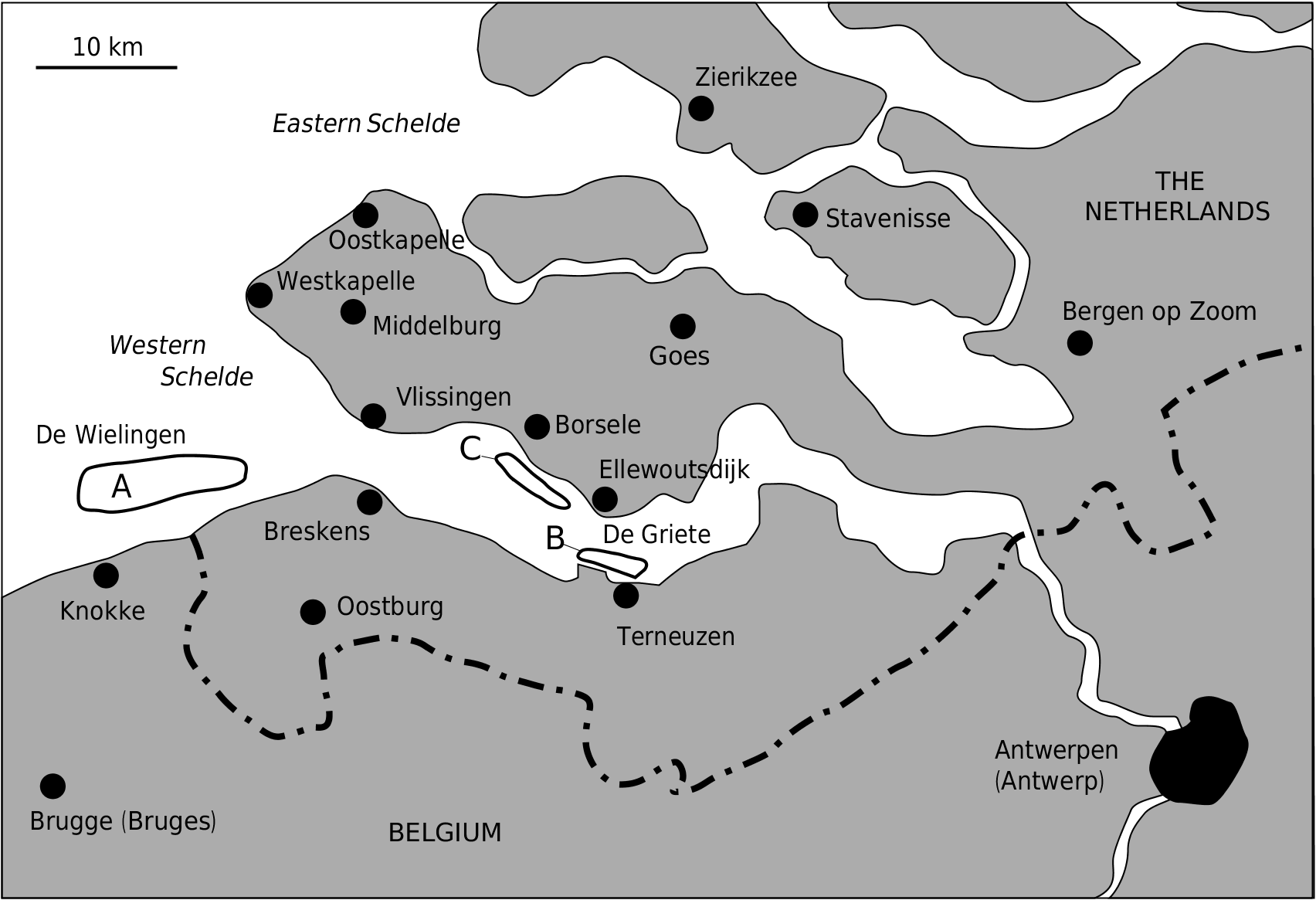
TYPE LOCALITY. — Scheldt river estuary, The Netherlands ( Fig. 1 View FIG ).
DIFFERENTIAL DIAGNOSIS. — Diunatans luctoretemergo n. gen., n. sp. is a member of the family Balaenopteridae because it shows: 1) abruptly depressed supraorbital process of the frontal; 2) lateral border of the supraoccipital overhanging the temporal fossa; 3) prominent squamosal crest; 4) parietals overlapping the posteromedial corner of the supraorbital process of the frontal; 5) ascending process of the premaxilla reaching the nasal; 6) maxilla, premaxilla and nasal reaching frontal and/or supraoccipital in a rectilinear or almost rectilinear line; 7) tympanic bulla with a weak, single and simple main ridge; and 8) anterior process of periotic triangular and ventromedial-dorsolaterally compressed.
Diunatans luctoretemergo n. gen., n. sp. differs from: – all extant Balaenopterinae , extant Megaptera novaeanglia (Borowski, 1781) and the nominal fossil rorquals B. cortesii ; B. cuvierii (Fisher, 1829) ; Megaptera hubachi Dathe, 1983 and Megaptera miocaena Kellogg, 1922 , in the morphology of the squamosal, the large, robust and prominent occipital condyle, the size of the tympanic bulla (relatively the largest of the family) and the orientation of its sigmoid process, and in the prominent promontorial groove of the periotic;
– Balaenoptera siberi , in the morphology of the squamosal, the large, robust and prominent occipital condyle, in the size of the tympanic bulla and the direction of its sigmoid process, and in the rounded pars cochlearis (pers. obs.). This species is poorly described. It shows basal features and does not belong in the genus Balaenoptera View in CoL (Bisconti pers. comm.; pers. obs.);
– fossil B. cortesii var. portisi Sacco, 1890 ; B. ryani (Hanna & McLellan, 1924) ; Cetotheriophanes capellinii ; Parabalaenoptera baulinensis Zeigler, Chan & Barnes, 1997 , by the short rectangular nasal;
– fossil B. davidsonii (Cope, 1872) because this species is based on a few mandibular characters only and therefore – in fact – may not be considered a valid taxon.
The fossil Megapteropsis robusta Van Beneden, 1872 (= Megaptera affinis Van Beneden, 1880 ); B. borealina Van Beneden, 1880 ; B. definata ( Owen, 1844) ; B. emarginata ( Owen, 1844) ; B. gibbosa ( Owen, 1844) ; B. rostratella Van Beneden, 1880 ; B. sibbaldina Van Beneden, 1880 ; Plesiocetus goropii Van Beneden, 1859 (= B. musculoides Van Beneden, 1882 ) are considered nomina dubia (see Appendix 1) and are therefore not compared to D. luctoretemergo n. gen., n. sp.
Diunatans luctoretemergo n. gen., n. sp. is not compared to B. gastaldi (Strobel, 1875) because this is an eschrichtiid ( Bisconti 2008); nor to Eobalaenoptera harrisoni Dooley, Fraser & Luo, 2004 because this a mysticete of uncertain taxonomic affinity ( Deméré et al. 2005). Additionaly the nominal fossil rorquals B. cortesii , B. cortesii var. portisi and B. cuvierii are probably not balaenopterids and are awaiting taxonomic revision ( Deméré et al. 2005; Bisconti in litt.)
DESCRIPTION
The description is based on NHG 22279 and NHG 22347. Both small crania have a zygomatic width of c. 750 mm. Th ey belong to full grown individuals because all cranial sutures are fused, the posterior process of the periotic is firmly attached to the cranium, and NHG 22279 shows cervical vertebral epyphyses firmly fused to the vertebral bodies.
Skull
Maxilla. About 115 mm of the ascending process of the maxilla is preserved. In dorsal view it is broad posteriorly (42 mm), ventrodorsally flattened and interdigitated with the frontal on the vertex (as in Balaenoptera ). In anterior view, the cross-section is comma-shaped, with a medial thickening. In lateral view the slope of the ascending process of the maxilla and premaxilla is c. 15° to the horizontal plane ( Figs 2 View FIG ; 4 View FIG ).
Premaxilla. About 109 mm of the ascending process of the premaxilla is preserved. It is wedged between the nasal and the ascending process of the maxilla. Posteriorly it overlays the frontal, dorsally it is relatively wide and in anterior cross-section it is comma-shaped ( Figs 2 View FIG ; 4 View FIG ).
Nasal. The nasal is similar to the nasal of extant rorquals B. physalus (Linnaeus, 1758) , B. musculus (Linnaeus, 1758) and Megaptera novaeangliae , but differs from that of B.acutorostrata , B.omurai , B.edeni Anderson, 1878 and B. borealis Lesson, 1828 (M. Bisconti pers.comm.; pers. obs.),the nasals of which lack the dorso-medial crest and show a convex anterior side. Th e nasal is roughly trapezoidal in dorsal view, slightly narrowing posteriorly and the nasals are wedged between the ascending processes of the premaxillae. Posteriorly they contact the frontals on the vertex.The dorsal surface of the naso-frontal suture is damaged; however it seems that the naso-frontal contact was probably not thoroughly interdigitated. Anteriorly the nasals are (slightly) concave ( Figs 2 View FIG ; 3 View FIG ).
Frontal. The frontal is exposed over the entire width of the vertex (anteroposterior L = 20 mm laterodorsally on the vertex) and protrudes dorsomedially (which results in a chevron-like shape in dorsal view). In lateral view, it is exposed dorsally over at least 120 mm. It is overlapped anterodorsally by the ascending process of the maxilla, and by the parietal posteriorly. Th e lateral extensions of both supraorbital processes are missing; only their posteromedial bases are preserved.Th e parietal overlaps the posteromedial corner of the supraorbital process. The base of the supraorbital process of the frontal is abruptly depressed. On the anteromedial wall of the temporal fossa, the fronto-parietal (coronal) suture runs at an angle of 45° in the direction of the anterior end of the optic nerve conduct. It continues dorsal to the base of the supraorbital process, and contacts the alisphenoid posteroventral to the posterior base of the optic nerve conduct ( Figs 2 View FIG ; 4 View FIG ).
Parietal. Both parietals are completely preserved.The parietal is not exposed on the vertex, except for a 5 mm wide triangular lateral wing. Th e anterior process of the parietal is vertical to slightly concave in anteroposterior section, and reaches c. 50 mm anterior to the posterior end of the ascending process of the maxilla and c. 40 mm to that of the premaxilla.From the posterodorsal side of the alisphenoid, the parietalsquamosal suture starts.It extends posterodorsally upwards, to join the posterior side of the lateral border of the supraoccipital shield (lambdoid crest),just behind the posterior side of the optic nerve conduct (similar to sutures noted in B. musculus and M. novaeangliae ). The parietal is pressed against the lateral border of the supraoccipital, up to the fronto-parietal suture at the vertex. Th e dorsolateral parietal border strongly overhangs the temporal fossa ( Figs 2 View FIG ; 4 View FIG ).
Vomer. The vomer is visible anteriorly as a V-shaped gutter-like bone, lateroposteriorly covered by (fragments of) the palatine. In ventral view, the vomer is exposed between the palatines (however because of damage) and posterior to the palatines, where it is part of the medioventral wall of the choanae extending posteriorly till the anterior side of the bop. Posteriorly it covers the basicranium and posterolaterally the pterygoids ( Figs 3 View FIG ; 4 View FIG ).
B
22279-a, b: A, right lateral view, line drawing; B, right lateral view,
Palatine. In ventral view, the palatine extends posterolaterally almost to the level of the dorsolateral edges. Posterodorsally the palatine contacts the anteroventral wall of the optic nerve conduct of the frontal ( Figs 3 View FIG ; 4 View FIG ).
Pterygoid. A small part of the pterygoid contributes to the anteromedial edge of the foramen pseudovale. Most of the wall of the pterygoid sinus is made up by the pterygoid; only its posterior part is formed by the basioccipital. Medioventrally the medial wing of the pterygoid sinus extends as far posterior as to the centre of the pars cochlearis, where it contacts the descending process of the basioccipital. The pterygoid fossa is wide compared to the zygomatic width of the skull (c. 110 mm in transverse diameter). Laterally the pterygoid is overlaid by the squamosal. In anterolateral view, the pterygoid is ventrodorsally elevated and forms part of the ventral wall of the optic nerve conduct (as in B. acutorostrata ) ( Figs 3 View FIG ; 4 View FIG ; 6 View FIG ).
Alisphenoid. The alisphenoid shows a triangular form. It is narrow, and positioned a few centimetres posteroventral to the posterior base of the optic nerve conduct ( Fig. 2 View FIG ).
Squamosal. In lateral view, the squamosal is anteroposteriorely short and not pointed, wide in posterior and anterior views, and is distinctly separated from the supraoccipital. It is large and especially strongly developed towards the lateral border of the supraoccipital shield. Th e posterodorsal side of the squamosal shows a bulge. Th e zygomatic process is slender. It is tapering anteriorly; the most anterior part is missing. Th e zygomatic process seems to diverge (slightly) from the skull axis, but due to damage, its orientation is not clear. The mediodorsal edge of the zygomatic process possesses a sharp sigmoid crest that merges posteriorly with the lambdoid crest. The dorsal edge of the zygomatic process is downwards sloping (c. 20° to the horizontal plane). Th e postglenoid process is thick, rounded ventrally and oriented posteroventrally at an angle of c. 100° to the horizontal plane. It tapers medially, becomes blade-like, and is transversely oriented. Th e articular surface for the mandibular condyle is concave and narrow. The postglenoid fossa (external auditory meatus) is moderately wide anteroposteriorly, with a wide, spoon-shaped depression medially. The postglenoid fossa is not high but quite wide. The squamosal cleft runs parallel to the parietal-squamosal suture. It is long and slen- der, with a pointed apex located far posteriorly. In dorsal view the squamosal cleft is completely visible (contrary to the condition in B. acutorostrata ) and similar to the condition noted in M. novaeangliae . The foramen pseudovale is mainly formed by the squamosal, and only the medial anteroventral part is covered by the pterygoid. The anteroventral squamosal-pterygoid suture is strongly concave. The squamosal-parietal suture is medially straight. The posterior wall of the temporal fossa is medially convex and bulges into the temporal fossa. The somewhat roughly surfaced sternomastoid fossa is a shallow depression, which is ventromedially restricted by a weak crest ( Figs 2-7 View FIG View FIG View FIG ).
Supraoccipital. The sub-triangular supraoccipital shield shows a truncated, rectilinear apex; it is strongly concave medially and lacks a medial crest. In dorsal view the lateral border of the supraoccipital is concave and strongly overhangs the temporal fossa ( Figs 2 View FIG ; 4 View FIG ; 5; 6C View FIG ; 7 View FIG ).
Basioccipital. The basioccipital is wide mediolaterally and anteroposteriorly short. Anteroventrally the basioccipital is covered by the vomer. The basioccipital descending process is strong, mediolaterally wide and anteroposteriorly short, as in B. musculus ( Figs 3 View FIG ; 5-7).
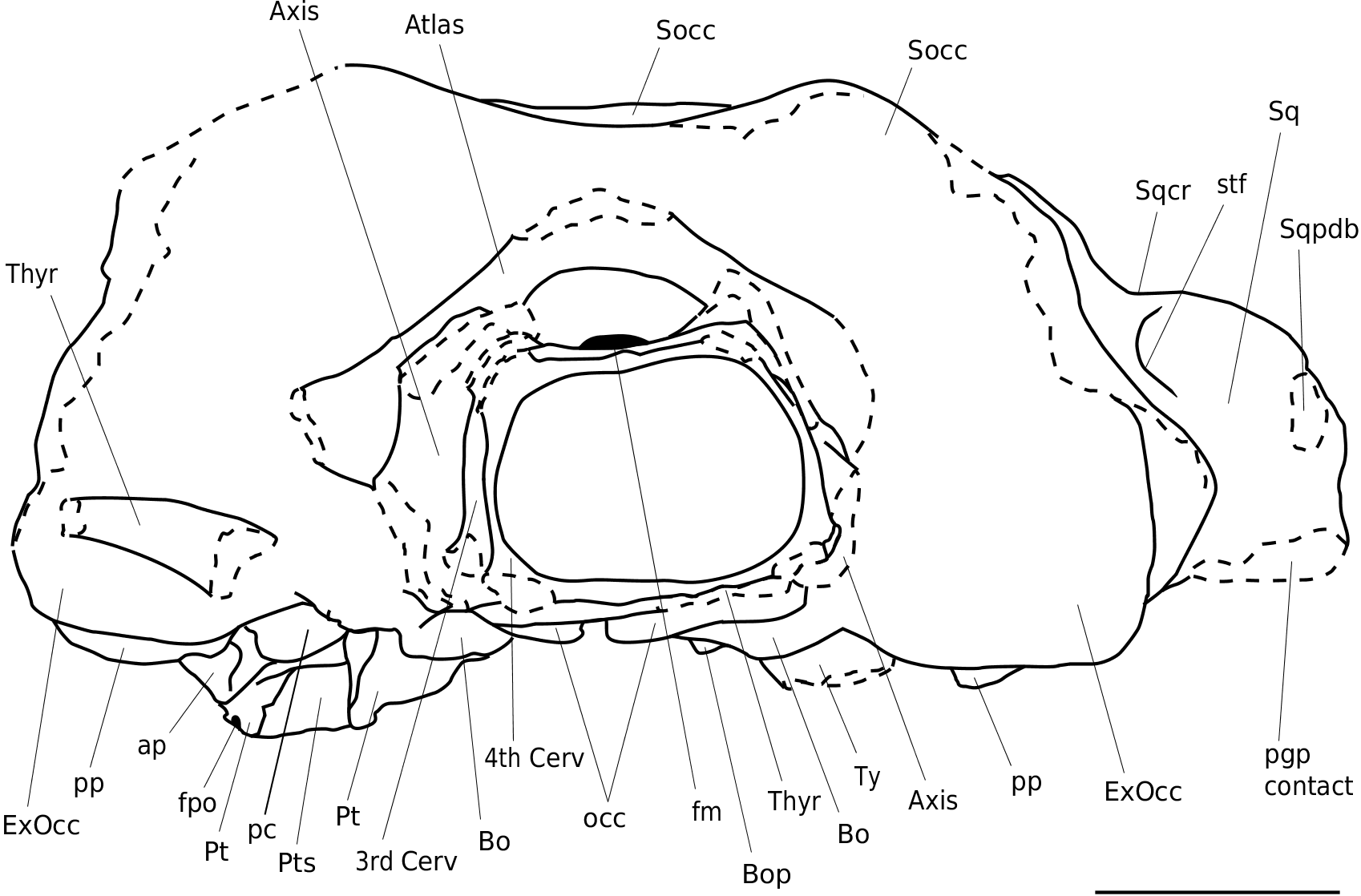
Exoccipital. The occipital condyles are massive, robust and much more protruding than the rather weak and rounded condyles of the extant Balaenopterinae but – to some extent – more similar to the condyles of M. novaeangliae . The well-developed occipital condyle is protruding strongly and extends about as far posteriorly as the exoccipital (in lateral view the condyles are clearly visible, while in all extant rorquals the condyles are partly or completely hidden behind the lateral side of the exoccipital and the occipital). Th e exoccipital is strong and wide and is oriented posteromedially ( Figs 2-7 View FIG View FIG View FIG ).
A Socc
fm
Sqcr Sq Sqpdb occ bo pc bop pp ExOcc eam
pgp
B
22347-a: A, posterior view, line drawing; B, posterior view, photo-
Petrotympanic
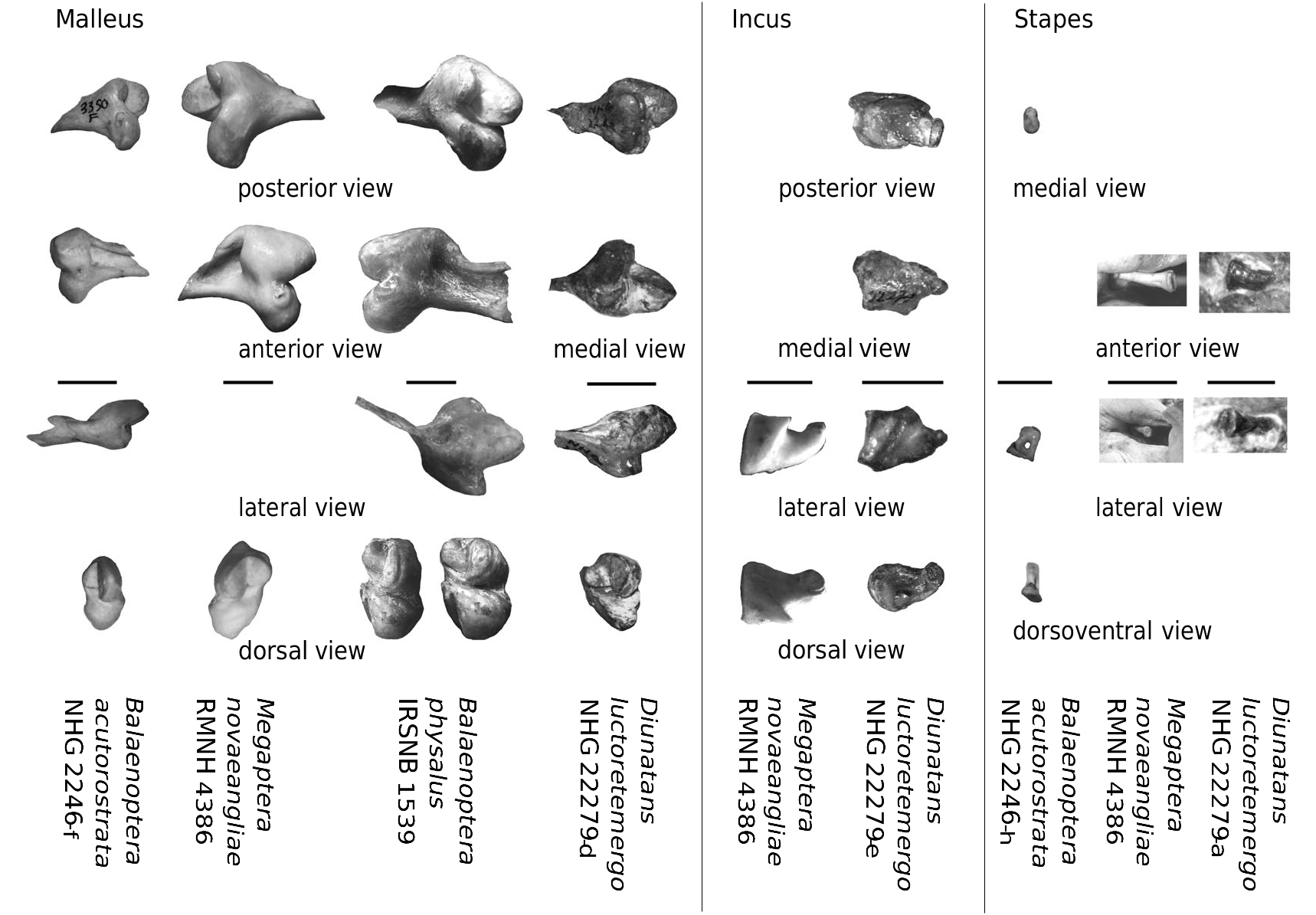
Tympanic bulla. After the removal of the left tympanic of the holotype (NHG 22279), the ventral face of the tympanic has been prepared, leaving the maleus and the incus nicely exposed. The greatest mediolateral width is at the level of the sigmoid process, while the greatest length runs parallel to the axis of the cranium. Anteriorly the lip is flattened and smooth. The anterior opening of the Eustachian tube is wide mediolaterally and much wider than in extant rorquals. The anterior half of the involucrum is depressed. It has a few rather weak transverse striae in this region; otherwise the surface is smooth. Anterolaterally, lateral to the Eustachian opening, the involucrum is mediolaterally wide. Th e involucrum is posteriorly wider, where it is also dorsoventrally flattened (as in all Balaenopteridae ) and has a weak dorsal lobe. Th e dorsomedial ridge is weak. The medial keel is weak and broad. Th e ventral lobes are not preserved. Th e sigmoid process is roughly perpendicular to the long axis of the tympanic, whereas it is inclined 17° to this axis in extant Balaenopteridae . Th e sigmoid process is well developed and is limited anteriorly by a wide sulcus. The conical apophysis is separated from the sigmoid process by a wide sub-triangular sulcus. Th e malleus is fused to the anterior side of the sigmoid process. The posterior pedicle of the tympanic is oriented anterolaterally-posteromedially. It is relatively short, wide, strong and located far posteriorly. The bulla is the largest known bulla of the family (relative to the zygomatic width of the cranium) and is wide compared to its length (length/width ratio: 1.24) ( Figs 3 View FIG ; 4 View FIG ; 7-9 View FIG View FIG View FIG ).
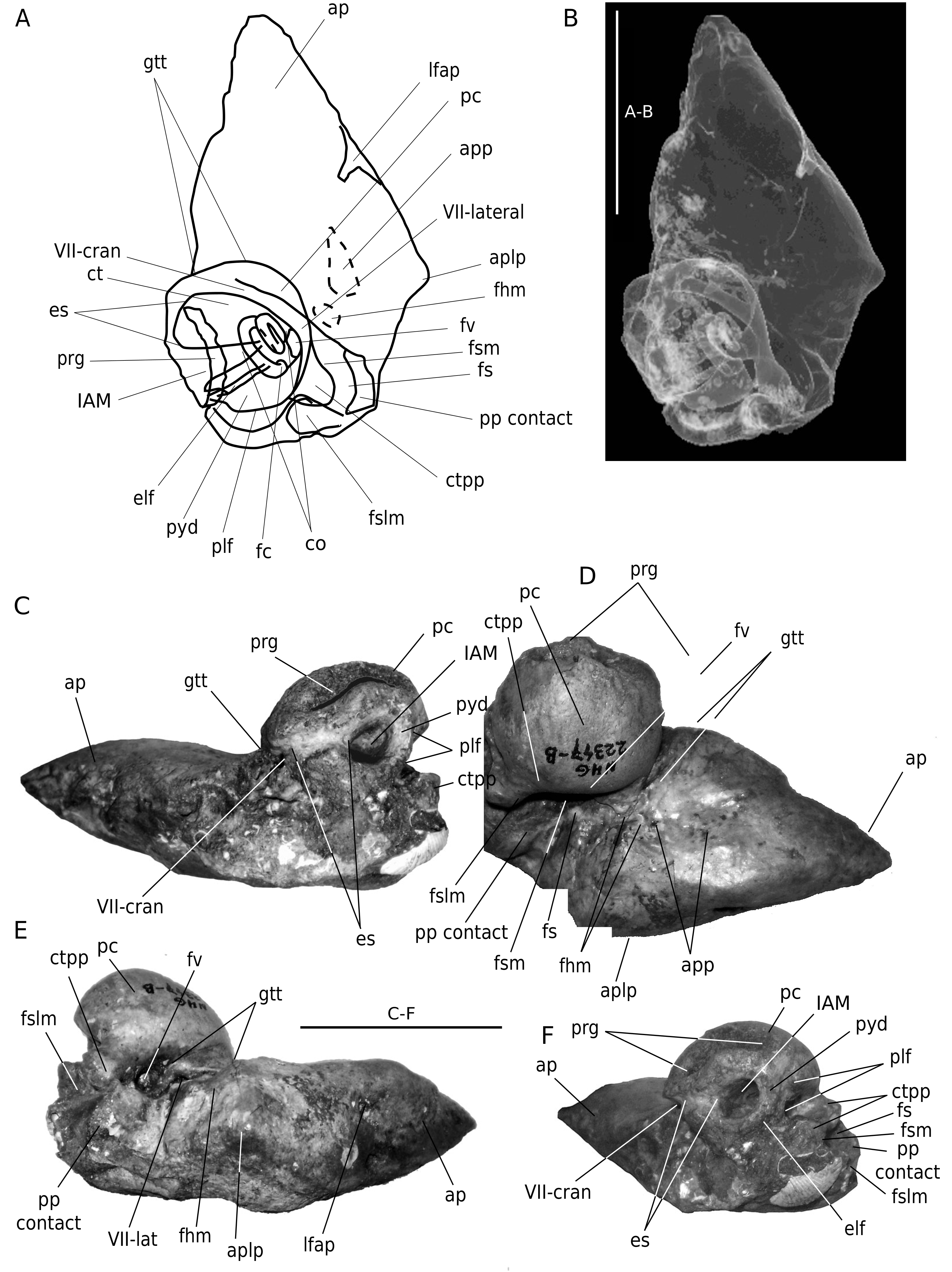
Periotic. The dorsolateral,dorsal and the dorsomedial (endocranial) side of the periotic of the holotype is still embedded in sediment and is not available for study.Descriptions of these views are therefore based on the paratype (NHG 22347). The left periotic of the paratype was removed from the skull, but the posterior process remains in situ.
On the holotype the anterior process of the periotic, in ventromedial view, is big and almost equilaterally triangular. It is almost twice as long as the anteroposterior length of the pars cochlearis. The anterior process is thick,ventromedially-dorsolaterally flattened and rounded at its ventrolateral and mediodorsal edges, similar to the anterior process of B. physalus , but shorter, wider and more ventromedially-dorsolaterally flattened. Both the ventrolateral and the mediodorsal sides of the anterior process are straight to slightly convex and smooth. Th e anterior half of the anterior process is covered by the squamosal when viewed ventromedially. In ventromedial view, the pars cochlearis contacts the anterior process over a very short distance only (20 mm). Th is is due to the short pars cochlearis and the fact that the endocranial facial sulcus is not roofed medially. The transverse elongation of the pars cochlearis is strongly restricted, and the roof of the medial expansion of the endocranial facial sulcus is apparently not yet fully ossified. In ventromedial view the pars cochlearis is almost circular and strongly bulbous. Such a bulbous pars cochlearis is not yet encountered in fossil and extant Balaenopteridae . Only extant Megaptera seem to show a similar (but weaker) feature. The pars cochlearis in Balaenopteridae is usually much more elongate, rather than sub-circular and globose. Th e transverse elongation of the pars cochlearis is limited in D. luctoretemergo n. gen., n. sp. (L/W ratio: 1.26 [holotype] to 1.23 [paratype]). The caudal tympanic process is strongly developed, obtuse, rounded and tube-shaped, Posteromedially it expands into a shelf. Th e anteroposterior distance between the anterior process and the posterior process of the periotic is large, and filled by the medially spoon-shaped and anteroposteriorly wide postglenoid fossa. Anteriorly, and separating the pars cochlearis from the anterior process, is a weak groove for the tensor tympani muscle, as still present in some extant rorqual specimens.Halfway on the ventrolateral side (ventrolateral ridge sensu Geisler & Luo 1996) is a large and deep foramen. Th e suprameatal fossa (squamosal fossa sensu Geisler & Luo 1996) is weak at the edges. Posteriorly, on the ventrolateral ridge is a rather prominent subtriangular ventrolateral projection (aplp sensu Geisler & Luo 1996). Medial to the ventrolateral projection is the anteroposteriorly oriented anterior pedicle of the periotic. This pedicle is triangular and short. Posterior to this pedicle, lateral to the pars cochlearis is a small shallow and hardly visible fossa for the head of the malleus. Lateroposterior to the mallear fossa is a deep and wide depression, therefore the periotic lacks a fossa incudis. Th e incus makes contact only to a narrow mediolaterally oriented crest, just posterior to the mallear fossa. Posterior to this depression is the rather slender, flattened crista petrosa:the base of the posterior process of the periotic. Medial to the crista petrosa is the stapedial muscle fossa. It is deep wide and well defined dorsally.Dorsomedial to the lateral facial sulcus is the stylomastoid fossa. It is also deep, short, and well separated from the stapedial muscle and facial nerve fossae by a thin transverse septum. The fenestra cochlearis is large (diameter> 6 mm) and subcircular. Th e lateroposterior opening of the facial nerve canal is circular and narrow (diameter c. 2 mm). The posterior process of the petrosal is massive and is firmly attached to the cranium. It is fairly short mediolaterally and anteroposteriorly flattened. Ventrodorsally it is high. Th e posterior process is at a right angle to the axis of the anterior process.
22279-a, b, posterior view of cranium and vertebrae, line drawing.
On the periotic of the paratype, the central area of the dorsolateral side is still covered by a thin layer of sediment and the surface of the bone is smooth at the ventrolateral and mediodorsal border, but probably slightly more rugose at the central area, where the sediment still partly covers the bone. In mediodorsal view, the internal acoustic meatus is sub-circular to oval and is isolated anteriorly from the facial nerve by a thick, robust (14 mm) crista transversa, and posteriorly from the perilymphatic foramen by the pyramidal process. The posterior wall of the perilymphatic foramen is not ossified. The endolymphatic foramen, just ventral to the posterior side of the internal acoustic meatus, is damaged. Th e endocranial facial foramen (VII-cran) is either damaged, or positioned very laterally. It is very small and circular (3 mm). Th e facial nerve canal continues over the mediodorsal side of the crista transversa, towards the acoustic meatus, to the extent that the endocranial foramen for the facial nerve and the internal acoustic meatus are interconnected by a deep, long and narrow sulcus, running over the crista transversa in anteroposterior direction. Ventral to the internal acoustic meatus is a prominent promontorial groove, similar to the structure we observed in some of the periotics of more archaic Balaenopteridae from the Antwerp Lower Pliocene Kattendijk Sands (Bisconti et al. unpubl. data) and from the Lee Creek Mine , North Carolina, USA (pers. obs.). This structure is absent in extant rorquals ( Figs 3-8 View FIG View FIG ; 10 View FIG ; 11) .
Malleus. The description is based on the left malleus of NHG 22279 (holotype). The malleus, incus and stapes were preserved in articulation. The pedicle of the malleus is fused to the anterior edge of the sigmoid process of the bulla. It is robust and bears two parallel columns separated by a deep groove. On its ventral side, the head bears two semi-circular articular facets for the incus. Th ey are at an angle of c. 90°. Th e dorsal facet is approximately twice as large as the ventral one. Th is facet is long compared to that of B. acutorostrata , but similar in size to that of B. physalus and M. novaeangliae and therefore long compared to the skull size. Both facets are slightly convex ( Figs 8 View FIG ; 11; 12 View FIG ).
Incus. The incus is sub-triangular in medial view. It is 13 mm long and tapers from 8.5 mm anteriorly to 2.7 mm posteriorly. It has an obtuse crus breve. The anterior contact surface to the malleus is big: 8 × 8.5 mm ( Figs 8 View FIG ; 11; 12 View FIG ).
Stapes. The stapes is very long and slender(L: 9.5 mm) and very slightly curved, as in M. novaeangliae . It has a diameter of 1.7 mm in the middle, widening to 3.2 mm posterolaterally ( Figs 8 View FIG ; 11; 12 View FIG ).
Post-cranial skeleton
Thyrohyal. On the left posterior side of the supraoccipital a c. 101 mm long and rounded fragment of a hyoid is connected by matrix. Most probably this is a part of the thyrohyal. Its thickest part (48 mm diameter) is located towards the midline of the cranium. On the right side a 80 mm bone fragment sticks to the ventral side of the atlas. Th is could be a part of the right thyrohyal ( Figs 2-4 View FIG View FIG View FIG ; 7 View FIG ).
malleus, incus and stapes tympanic bulla periotic
Cervical vertebrae. Four articulated cervical vertebrae are preserved. Th e atlas is well preserved and is 250 mm wide. Its thickest part is located on the centre of the vertebral body (45 mm); more lateral the thickness gradually reduces to about 35 mm. The neurapophysis is low and carries long, fairly slender and pointed transverse processes. The ventral surface of the vertebral body is almost horizontal. The condylar facets are not visible because the four cervical vertebrae are still attached to the cranium by sediment. The strong curvature visible at some uncovered parts, and the strong curvature of the occipital condyle of the paratype (NHG 22347), hint to a strongly arched surface. Th e vertebral body of the axis, C3 and C4 are 150, 145 and 130 mm wide respectively, with their largest thickness at the centre (28, 23 and 19 mm, measured ventrally).They all show a prominent and sharp medial keel on the dorsal surface of the vertebral body. Th e lateral sides of the vertebral body are more posteriorly located than the central parts, resulting in a weak V-shaped outline in dorsal view. Th e transverse processes of these vertebrae are missing ; only the basal joints are preserved. Th e epiphyses show almost no trace of sutures and are well connected to the vertebral body ( Figs 2-4 View FIG View FIG View FIG ; 7 View FIG ).
STRATIGRAPHY
The holotype and paratype specimens were trawled from an unknown locality in the Westerschelde , The Netherlands in 1938 ( Fig. 1 View FIG ). Late Miocene-Mid Pliocene cetacean fossils have been trawled from three locations in the 1930s: De Wielingen, in front of Knokke ( Fig. 1A View FIG ) ; De Griete and De Braakman near Terneuzen ( Fig. 1B View FIG ) and along the riverbank near Borsele and Ellewoudsdijk ( Fig. 1C View FIG ).
The damaged edges of the holotype skull reveal that it must have been fairly complete and that rostrum, frontals and postcrania must have been damaged during dredging. Th e absence of wear caused by reworking or secondary transportation, the presence of both petrotympanic complexes, and the position of the cervical vertebrae in articulation, indicate that the skeleton was fairly completely preserved, on or near the river bed.
Miocene and Pliocene fossils of marine mammals, as well as (to a lesser extent) Late Pleistocene fossils of terrestrial and marine mammals, are known from the Westerschelde ( Drees 1986; Ebbing et al. 1993). Th e Miocene and Pliocene fossils originate from the Antwerpen Sand Member (Middle Miocene), the Deurne Sand Member (Late Miocene), the Kattendijk Sand Member (Early Pliocene), the Luchtbal Sand Member (late Early Pliocene) and the Oorderen Sand Member (Middle Pliocene) and are reworked from these strata by the eroding river Schelde ( Drees 1986; Ebbing et al. 1993).
Parts of the original matrix in which the holotype skull was embedded, are still attached to the cranium and between the vertebrae. From this cemented, coarse and sandy matrix seven species of molluscs were collected (Appendix 4). Three molluscs ( Calyptrea chinensis (Linnaeus, 1758) , Arctica islandica (Linnaeus 1767) , and Corbula gibba gibba (Olivi, 1792)) occur from the Miocene onwards, and therefore do not provide specific stratigraphic information ( Marquet 1998, 2005). Parvicardium scabrum (Philippi, 1844) also encompasses a wide stratigraphic timeframe (Pliocene to Recent), but is not known from Miocene strata ( Marquet 2005). Ringicula buccinea (Brocchi, 1814) is of Miocene to Early Pliocene occurrence and the sample strongly resembles the more archaic morphology of the species. Compared to Middle and Late Pliocene specimens, it is smaller, has a narrower outer lip and a more shallow suture. Th is morphology also occurs in the Kattendijk Formation (Early Pliocene) at Schijnpoort, Antwerpen, Belgium ( Marquet 1998). Middle Pliocene or younger records of this species in the North Sea basin are not known. Isocrassina omalii (de la Jonkaire, 1823) is a bivalve which is typical for the Early Pliocene (Kattendijk and Luchtbal Sand Members) and scarce finds are known from the basal crags of the Middle Pliocene (Oorderen Sand Member) but seem reworked ( Marquet 2005). Thracia altenai Glibert & van de Poel, 1966 occurs from the Early to the Middle Pliocene ( Marquet 2005). Th e absence of Palliolum gerardi (Nyst, 1835) seems noteworthy since this species is extremely common in the late Early Pliocene Luchtbal Sand Member.
Taking all evidence into account, an Early Pliocene (Zanclean; between 5.0 Ma and 4.7-4.4 Ma; Louwye et al. 2004) origin of this cranium is assumed.
In the Schelde estuary, Early Pliocene strata were deposited in a near coastal environment with a water depth of probably less than 25 m because fossil Balanus species are common on Antwerp cetacean bones from the Kattendijk sands (pers. obs.). Seasurface temperatures were cool to temperate (less than 7.2°C for April sea-surface temperatures) ( Gaemers 1988; Louwye et al. 2004; Lambert 2007).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Diunatans luctoretemergo
| Bosselaers, Mark & Post, Klaas 2010 |
Diunatans luctoretemergo
| Bosselaers & Post 2010 |
Diunatans luctoretemergo
| Bosselaers & Post 2010 |
Eobalaenoptera harrisoni
| Dooley, Fraser & Luo 2004 |
Balaenoptera siberi
| Pilleri & Pilleri 1989 |
Megaptera hubachi
| Dathe 1983 |
Megaptera miocaena
| Kellogg 1922 |
B. cortesii var. portisi
| Sacco 1890 |
Balaenopterinae
| Flower 1865 |
Balaenoptera
| Lacepede 1804 |