
Discotettix (Discotettix) belzebuth ( Serville, 1838 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5217.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:86CD1EDF-8C38-4A90-888A-185B8481A6ED |
|
DOI |
https://doi.org/10.5281/zenodo.7409399 |
|
persistent identifier |
https://treatment.plazi.org/id/0129163A-B12C-6000-FCCA-FA1AFE58FD19 |
|
treatment provided by |
Plazi |
|
scientific name |
Discotettix (Discotettix) belzebuth ( Serville, 1838 ) |
| status |
|
Discotettix (Discotettix) belzebuth ( Serville, 1838) View in CoL View at ENA ( Figs 5–14 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 View FIGURE 14 )
Vernacular name: Bornean Spiky Pygmy Devil
Tetrix belzebuth Serville, 1838: 759 View in CoL [original description, type locality: originally Java but actually probably Borneo ].
Tettix belzebuth: Stål 1873: 152 [listed in the catalog].
Discotettix belzebuth: Bolívar 1887: 306 View in CoL [included in the revision]; Rehn, 1904: 670 [new records]; Hancock 1907a: 6 [included in the revision]; Hancock 1907b: 213 [new records]; Kirby 1910: 2 [included in the catalog]; Hancock 1913: 39 [new records]; Willemse 1930: 8 [new records]; Günther 1938: 301 [included in the revision]; Steinmann 1970: 216 [listed in the catalog]; Blackith 1992: 46 [listed in the catalog]; Yin et al. 1996: 866 [listed in the catalog], Otte 1997: 32 [listed in the catalog], Kočárek et al. 2015: 289 View Cited Treatment [new records, data on variability]; Kuřavova et al. 2017: 120–128 [data on the antennae morphology]; Tan & Wahab 2018: 123 View Cited Treatment [new records in Brunei Darussalam].
Discotettix armatus Costa, 1864: 59 View in CoL [original description, type locality: Borneo]; synonymized by Bolívar (1887).
Discotettix adenanii Mahmood et al., 2007: 1276 View in CoL [original description, type locality: Borneo: Kuching]; synonymized by Kočárek et al. (2015).
Type locality. According to the original description the type locality is Java, but since no locality label is present under the holotype (the only found specimen originating from Serville's collection in MNHN), we believe that the specimen originates from Borneo, from where the majority of the records of this species are. In MNCN, there are four specimens labeled ‘Java’ so it is not fully clear whether this species is/was present on Java. Borneo Island is the type locality of D. armatus , which is the type species of the genus Discotettix (a junior synonym of D. belzebuth ). The type specimen of D. belzebuth was considered lost, but was found in March 2016 by JS in MNHN. For D. armatus , we believe that the specimen still exists in the Naples collection ( Italy), but we were not able to contact the Museum or to get information on Costa's collection.
Material examined.
Type material. HOLOTYPE of D. belzebuth 1♀ (locality, date and collector labels missing), red label 'TYPE' and Günther's label ' Discotettix belzebuth Serv. K. Günther det.' present ( MNHN); PARATYPE of D. adenanii 1♂ Malaysia: Sarawak: “Adanan Bukra”, Gunung Serapi 3.VI.1988. ( UKM) (according to Kočárek et al. (2015) the holotype and the rest of the type series has probably been destroyed).
Additional museum material. 2♀♀, 1♂ Borneo : Malaysia: Sarawak: Mattang Collector Frivaldsky, det. J. Skejo ( MNCN) ; 1♀, 2♂♂ Borneo : Malaysia: Sarawak Collector Mjöberg, det. J. Tumbrinck ( NHRS) ; 1♀ Borneo: Malaysia: Sarawak: Bidi 1908. Collector. C. J. Brooks, det. J. Tumbrinck ( NCM) ; 1♂ Borneo : Malaysia: [Sarawak, Bidi] Collector: C. J. Brooks, det. J. Tumbrinck ( NCM) ; 1 ♀ Borneo: Malaysia: Sarawak: 75 km S of Miri Town, Niah Nat. Park [100 m a.s.l., forest around Niah Great Cave] 30.III.2012. Collectors A.V. Gorochov and M. Berezin, det. J. Skejo et T. Pushkar ( ZISP) ; 2♀♀ Borneo : Malaysia: Sabah: Kina-Balu-Gebirge Collector: Waterstradt, det. Hancock ( AMS) ; 1♀ Borneo: Malaysia: Sabah: Kinabalu NP: Poring [forest clearing] 8.VIII.1984. Collector S. Ingrisch, det. S. Ingrisch ( CJT) ; 2♀♀, 2♂ ♂ Malaysia: Sabah [labeled Nord-Borneo Collector Waterstradt det. J. Skejo ( MNCN) ; 3♂♂ Borneo: Malaysia: Sabah: Pajau River , Collector Mjöberg, det. J. Tumbrinck ( NHRS) ; 1♂ Borneo: Malaysia: Sabah: Kajan River Collector Mjöberg, det. J. Tumbrinck ( NHRS) ; 1♀, 2♂♂ Malaysia: Sabah [on original label written North Borneo] Collector: Watterstradt, det. K. Günther ( SMTD) ; 1♀ Borneo: Malaysia: Sabah: Batu Niah 4.VIII.1984. Collector S. Ingrisch, det. S. Ingrisch ( ZFMK) ; 1♀ Borneo: Malaysia: Sabah: Kinabalu NP: Poring: Bergil 10.IV.1997. [6º5'N, 116º33'E] [collected by fogging] Collector A. Floren, det. J. Tumbrinck ( ZFMK) GoogleMaps ; 1 ♀ Malaysia: Sabah: Crocket Range , 80 km S of Kota Kinabalu City [environment of the village Ula Kumanis at 800 m a.s.l.] 5 – 10.V.2006. Collector A. Sochivko, det. J. Skejo et T. Pushkar ( ZISP) ; 1♀, 3♂ Borneo: Malaysia: Sabah: [original labels „ Nord Borneo , Kina-Balu-Gebirge [at 1500 m a.s.l.] Collector Waterstradt, det. Brunner von Wattenwyl ( ZISP) ; 1♀ Malaysia: Sabah: Mt. Trus Madi: Tambunan distr [975 m a.s.l.] 25.IV–10.V.2006, Collector P. Udivichenko, det. J. Skejo et T. Pushkar ( ZISP) ; 4 ♂♂ Malaysia: Sabah: Mt. Trus Madi: Tambunan district [975 m a.s.l.] 25.IV–10.V.2006. Collector P. Udivichenko, det. J. Skejo et T. Pushkar ( ZISP) ; 2♀, 2♂ Borneo: Malaysia: Sabah: Mt. Trus Madi [1000 m a.s.l.] 13.–25.V.2007. Collector A.V. Gorochov, det. J. Skejo et T. Pushkar ( ZISP) ; 1 ♀ Borneo: Malaysia: Sabah: Mt. Trus Madi [1200 m a.s.l.] 13–24.I.2007. Collector A. Sochivko, det. J. Skejo et T. Pushkar ( ZISP) ; 1♂ label '99. 10064.' [without specified data] det. J. Skejo ( MNCN) ; 1♂ [without specified data] det. J. Skejo ( MNCN) ; 1♀, 1♂ Malaysia: Borneo [without specified other data] ( NMNH NASU) ; 2♀♀ Indonesia: Kalimatan: Kalimantan Timur [= East]: Marah 12.XI.1925. Collector: H. C. Siebers, det. K. Günther ( SMTD) ; 1 nymph (sex unteterminable) Indonesia: Kalimatan: West Kalimatan : Pontianak det. J. Skejo ( MNCN) ; 2♀♀, 2♂ ♂ Indonesia: Java [without other data specified] det. J. Skejo ( MNCN) ;
Additional material from online social media. All the records of D. belzebuth from iNaturalist may be found on the following link: https://www.inaturalist.org/observations?taxon_id=637240 (all records submitted by July 2022 are shown in Table 1 View TABLE 1 ); while for Flickr, ProjectNoah, and SpinelessWonders all the observations ( Figs 6–14 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 View FIGURE 14 ) are listed in the Table 1 View TABLE 1 .
TABLE 1. (Continued)
Distribution. The species is widespread in Borneo (many data from Malaysian and Brunei part, but few from the Indonesian part, probably due to the lack of research) and adjacent small islands (such as Labuan). This species was previously considered widely distributed on all the Greater Sunda Islands of the Malay Archipelago, namely on Borneo, Java, and Sumatra ( Serville 1838; De Haan 1843; Rehn 1904; Hancock 1907a; 1907b; Günther 1938; Mahmood et al. 2007; Kočárek et al. 2015; Tan & Wahab 2018). Distribution of the species in Java was/is possible, as in the Bolívar's collection in MNCN Madrid there are several specimens labeled ‘Java’, but its presence nowadays should be confirmed as there have been no records for more than a century. The species is not present in Sumatra ( De Haan 1843), with De Haan’s (1843) records likely belonging to D. selysi , which was not described at the time.
Taxonomic notes. Discotettix adenanii is considered a junior synonym of D. belzebuth . Seven well-developed pronotal projections ( Mahmood et al. 2007) are the diagnostic character of D. belzebuth as well. Günther (1938) pointed out the variability of this polymorphic species with certain specimens that are not easy to identify as D. belzebuth , but as a form with certain characteristics somewhat similar to Kraengia . The description of D. adenanii , on the other hand, completely fits D. belzebuth description and diagnosis. Variability of the species can be observed solely by comparison of Bolívar's drawing ( Hancock, 1907a: 6) with Günther's D. belzebuth figure (1938: p. 301, Fig. 2 View FIGURE 2 ). Kočárek et al. (2015) examined type specimen of D. adenanii and the variability of the pronotal morphology in D. belzebuth populations collected in different parts of Borneo, showing that all the studied specimens fit the variability of D. belzebuth . We agree with the conclusion of Kočárek et al. (2015) that D. adenanii is a pure synonym of D. belzebuth .
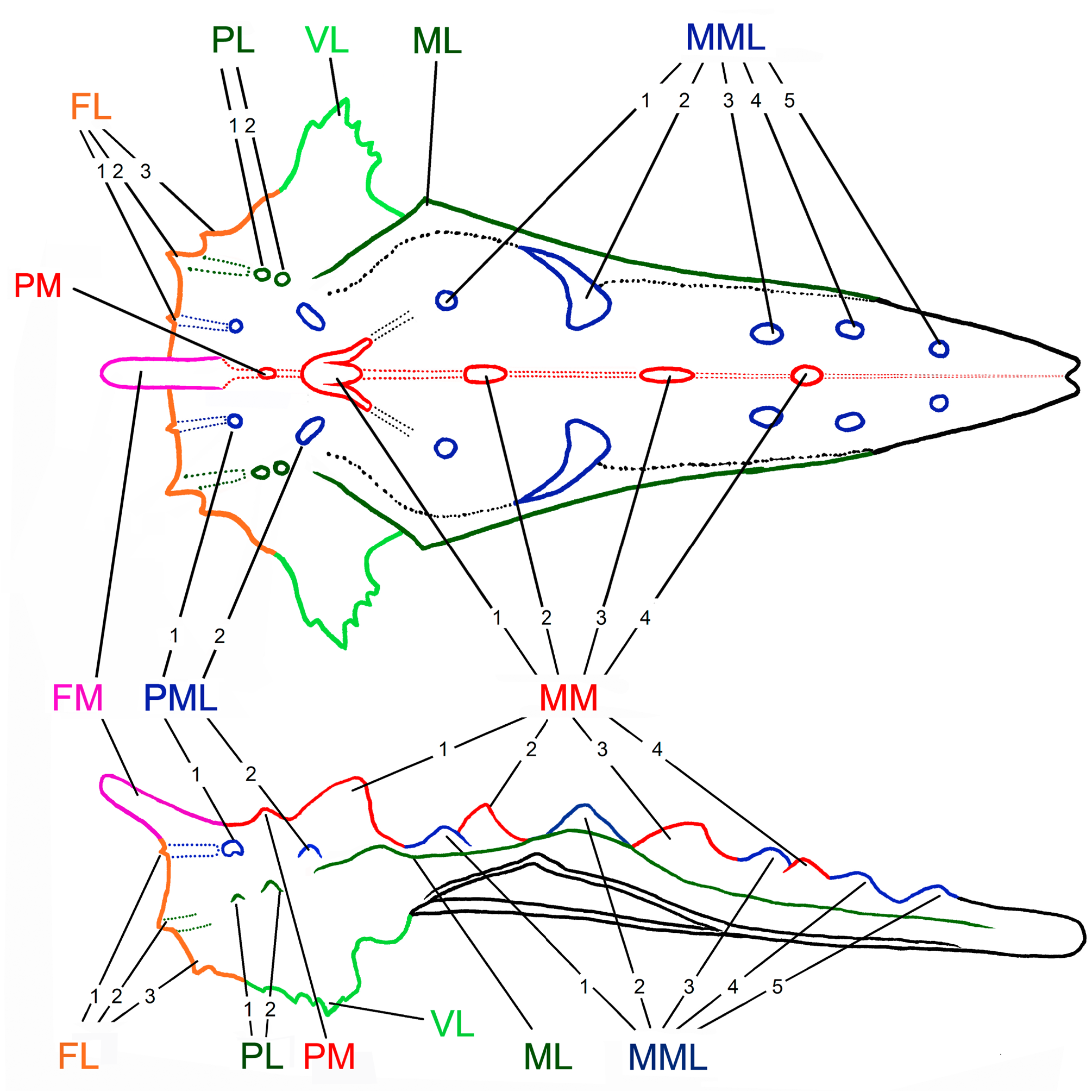
Diagnosis. This is the species with the highest (in lateral view) and spikiest/sharpest pronotal projections in the entire genus and by its general appearance it can readily be distinguished from all the other species of the genus. It is somewhat similar to D. kirscheyi Skejo, Pushkar, Tumbrinck et Tan sp. n. from NE Borneo, and to D. sumatrensis Skejo, Pushkar et Tumbrinck sp. n. from Sumatra. From D. scabridus the new species can be separated by the following characters: (I) bifurcation of the frontal costa between the eyes (on the lower margin of the compound eyes in D. scabridus ), (II) FM high and developed (present as a small tubercle in D. scabridus ), (III) MM elevated into spines (lower and saw-like in D. scabridus ), (IV) MML elevated into spines (present as low and triangular, compressed elevations in D. scabridus ), (V) ML strong and long, tooth-like (absent in D. scabridus ), (VI) interscapular area with parallel margins (wide and triangular in D. scabridus ). Discotettix belzebuth can be separated from D. selysi , D. doriae , and D. aruanus Skejo, Pushkar et Tumbrinck sp. n. by the following characters: (I) more than one well-developed protuberance on the pronotal disc, (II) long frontomedial projection of the anterior margin of the pronotum, (III) fore and mid femora are more slender, (IV) antennal segments are more specialized in morphology than in D. selysi , the widest and the most flattened is the 10 th segment (unlike the 9 th segment in D. selysi and D. doriae ). From D. sumatrensis Skejo, Pushkar et Tumbrinck sp. n. the species can be easily separated by the following characteristics: (I) larger body size, (II) all antennae segments are of black/dark color, (III) the 8 th antennal segment is the widest one (unlike the 7 th segment in D. sumatrensis Skejo, Pushkar et Tumbrinck sp. n.), (IV) the hind femur bears small lappets. D. kirscheyi Skejo, Pushkar, Tumbrinck et Tan sp. n. can be easily separated from D. belzebuth by the following set of characters: (I) in D. kirscheyi Skejo, Pushkar, Tumbrinck et Tan sp. n. antennae with more robust segments gradually become wider distally, not as specialized as in D. belzebuth, (II) D. kirscheyi Skejo, Pushkar, Tumbrinck et Tan sp. n. has short FM, not covering the whole head as in D. bellzebuth, (III) projections on the dorsal surface of the pronotum are smaller in D. kirscheyi Skejo, Pushkar, Tumbrinck et Tan sp. n., (IV) D. kirscheyi sp. n. has weak ML, (V) D. kirscheyi sp. n. has robust and serrated fore and mid femora, not as slender as in D. belzebuth and (VI) D. kirscheyi sp. n. has smaller body size.
Redescription.
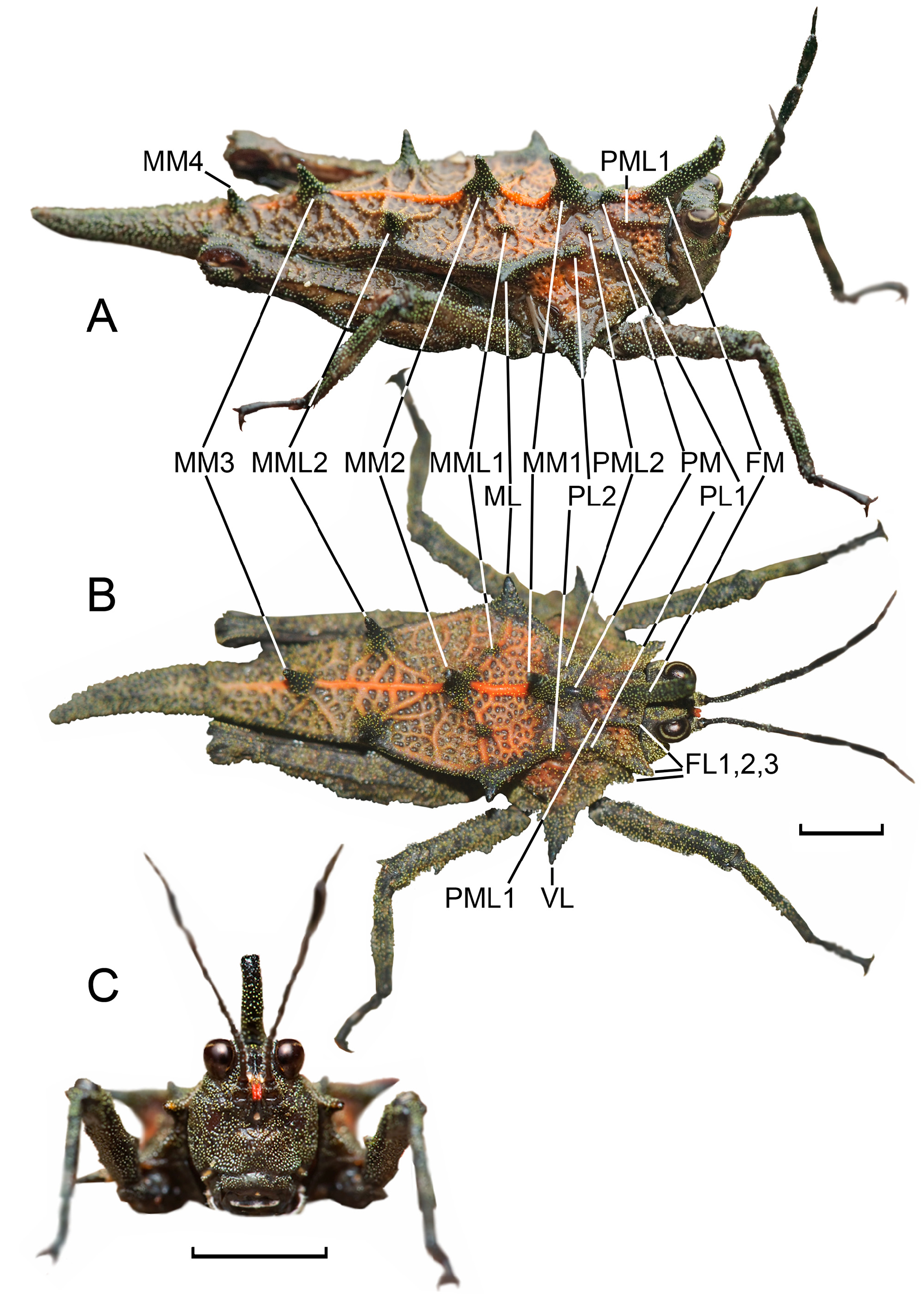
General characters. Medium to large sized, robust, species (13.01–17.02 mm), texture granulated, rugose; pronotum wrinkled, with numerous small tubercles, medium-sized and large protuberances on dorsal and lateral sides. Epizoic symbiotic bryophytes and algae are often present on the pronotal surfaces, so usually the specimens are characterized by cryptic colors ( Figs 8 View FIGURE 8 , 10 View FIGURE 10 ). Macropronotal.
Coloration. Coloration variable: from black and dark brown to brighter tints of brown: grayish, greenish, yellowish, reddish, somewhat purple; pronotal projections usually darker. The entire body, including antennae, may be of the same color, except brightly colored frontal carina on the head and the median carina of pronotum, while fore and mid tibiae and tarsi usually bear 1–3 lighter rings. Maxillary palpi black with pale colored joints between the segments ( Figs 5–14 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 View FIGURE 14 ).
Head. In dorsal and frontal view, vertex 2.21–2.76 times as wide as an eye. Fossula elliptic and deep.
The lower margin of the lateral ocelli a bit below the level of the lower margin of a compound eye. In frontal view, frontal costa narrow, bifurcated above lateral ocelli into subparallel, finely granulated facial carinae forming a very narrow scutellum. Scutellum slightly narrower than the antennal groove. Antennal groove below the lower margin of the compound eye ( Fig. 5C View FIGURE 5 ). Antenna with 13 antennomeres. A detailed description of the antennal morphometrics and morphology was published recently by Kuřavová et al. (2017). Antennal segments as follows: scapus (1 st antennomere) and pedicel (2 nd antennomere) massive; basal segments (3 rd to 6 th) elongated and circular in cross-section; central or subapical segments (7 th and 8 th) strongly widened, pennate, 8 th being the widest antennal segment (in holotype 2.22 times as long as wide); apical segment 9 th much smaller than the subapical and much larger than the rest of the apical segments; apical segments 10 th to 13 th reduced, very small, and borders between them barely visible. Antennomeres 3 rd to 9 th bearing saw-like margins, because of the presence of large basiconic sensilla ( Fig. 10 View FIGURE 10 ). When the body and the antennae are covered with algae and moss, the 8 th segment is always free of epizoic organisms ( Fig. 10 View FIGURE 10 ).
Pronotum. Pronotum wrinkled and granulated, covered in numerous small tubercles and larger projections. The posterior process of the pronotum slender, surpassing the hind knee for about half of the hind femur length. Disc of the pronotum at almost the same level along all length, and unlike other species of the genus, without a distinct depression behind the shoulder. Pronotum not descending backward. Morphology of the pronotal disc variable, usually with 4–7 unpaired projections of variable size on the median carina (FM, PM, and 2–5 MMs); 2–3 pairs of FL; 4–7 pairs of more or less distinct PMLs and MMLs; 3 pairs of PLs and MLs; and a pair of VL. Prozona short, subsquare. The anterior margin of the pronotum projected as a large digitate FM directed upwards and forwards above the head, covering the whole area of the fastigium of the vertex. FM often decurved with excised apex. Prozonal carinae slightly elevated, surpassing the anterior margin of the pronotum as dentiform FL1. Extralateral carinae elevated, surpassing anterior margin of pronotum as dentiform FL2. FL3 dentiform, small and weak, more distinct than FL1. Median carina extended along the whole length of the pronotum, tuberculated, but with smooth areas. Median carina bearing FM and 3–6 of large digitate medial projections well seen in profile. PM small, distinct in about a third of the examined specimens. MM1, MM2, MM3, and MM4 large and digitate (MM1<MM2>MM3>MM4). MM4 well visible in most of the specimens, but in some completely reduced. MM5 present as a spike in about every tenth specimen, but usually very small and almost invisible. Among mediolateral projections, PML1 is a small tubercle, almost indistinct; PML2 small; MML1 small; MML2 large; MML3 and MML4 distinct in a few examined specimens. PL1 and PL2 elongated, small, and almost indistinct. In the metazona humero-apical carinae forms a sharp humeral angle, projected outwards as strong spine-like or digitate ML. Interhumeral carinae hardly observable because of numerous net-like elevations and tubercles present in the whole disc. The apex of pronotum blunt, and shallowly excised. The lower part of the lateral lobe with finely serrate anterior and coarsely serrate posterior margin, elongated as VL of variable shapes from sharp spine-like to saw-like form, directed strongly outwards or in rare cases somewhat backward ( Fig. 5A, B View FIGURE 5 ).
Wings. The visible part of tegmen elongated and oval. Hind wing long, often not reaching the pronotal apex.
Legs. Femora more or less robust, and compressed laterally, but elongated in comparison to other species. The rough surface of the legs usually bears outgrowths and tubercles of variable size and sharpness. The dorsal and ventral margins serrate. Genicular teeth visible on all the knees, and additionally 1–3 weak teeth present on the dorsal and ventral margins of the fore and mid femora. Hind femur with small lappets on both dorsal and ventral margins. Lateral area of the hind femur with net-like elevations and weak carinae, ventro-external carina with teethlike outgrowths. Genicular teeth of the hind femora larger than the antegenicular. Both sides of the dorsal margin of the hind tibia finely serrated, additionally with 4–5 outer and 3–4 inner larger teeth.
Abdominal apex. Female subgenital plate with a triangular protrusion in the middle of the posterior margin. Ovipositor of variable shapes, usually elongated, but can be more robust, probably due to the ecological factors.
Measurements. BL ♂♂ 13.1–15.01 mm, ♀♀ 14.3–17.02 mm; PnL ♂♂ 16.04–19.14 mm, ♀♀ 18.5–22.34mm; PnW ♂♂ 7.99–8.24 mm, ♀♀ 9.04–9.66 mm; AnL ♂♂ 6.88–7.8 mm, ♀♀ 7.11–7.99 mm; TL ♂♂ 1.9–2.54 mm, ♀♀ 2.1–3.01 mm; TW ♂♂ 0.78–1.14 mm, ♀♀ 0.99–1.23 mm; fFL ♂♂ 3.29–4.46 mm, ♀♀ 4.1–5.16 mm; fFW ♂♂ 0.77–1.01 mm, ♀♀ 0.89–1.02 mm; mFL ♂♂ 3.49–4.72 mm, ♀♀ 4.2–5.09 mm; mFW ♂♂ 0.98–1.02 mm, ♀♀ 0.99–1.11 mm; hFL ♂♂ 6.99–10.03 mm, ♀♀ 8.1–10.28 mm; hFW ♂♂ 2.28–3.45 mm, ♀♀ 2.26–3.24 mm; OvL ♀♀ 1.38–2.11 mm; AnL/fFL ♂♂ 1.58–2.09, ♀♀ 1.55–1.71; VW ♂♂ 1.02–1.48 mm, ♀♀ 1.35–1.91 mm; EW ♂♂ 0.38–0.49 mm, ♀♀ 0.46–0.82 mm; VW/EW ♂♂ 2.21–2.52, ♀♀ 2.31–2.76; SW ♂♂ 0.31–0.37 mm, ♀♀ 0.32– 0.42 mm; AgW ♂♂ 0.19–0.34 mm, ♀♀ 0.29–0.41 mm; ScW ♂♂ 0.21–0.29 mm, ♀♀ 0.23–0.28 mm; SW/AgW ♂♂ 1.46–1.51, ♀♀ 1.41–1.56; SW/ScW ♂♂ 1.20–1.47, ♀♀ 1.21–1.35; As—L/W ♂♂ 2.41–3.1, ♀♀ 2.39–3.12; PrzW ♂♂ 3.35–3.59 mm, ♀♀ 4.59–4.82 mm; PrzL ♂♂ 3.7–4.11 mm, ♀♀ 4.29–4.99 mm; Prz—W/L ♂♂ 0.85–0.9, ♀♀ 0.84–0.91; TL/TW ♂♂ 2.85–3.17, ♀♀ 2.88–3.11; mFW/TW ♂♂ 0.81–1.29, ♀♀ 0.87–1.24; fFL/fFW ♂♂ 4.11– 5.41, ♀♀ 4.12–5.22; mFL/mFW ♂♂ 4.21–5.01, ♀♀ 4.28–4.91; hFL/hFW ♂♂ 3.01–3.67, ♀♀ 3.19–3.59; T1L/T3L ♂♂ 0.89–1.25, ♀♀ 0.9–1.11.









| MNHN |
Museum National d'Histoire Naturelle |
| UKM |
Universiti Kebangsaan Malaysia |
| MNCN |
Museo Nacional de Ciencias Naturales |
| NHRS |
Swedish Museum of Natural History, Entomology Collections |
| ZISP |
Zoological Institute, Russian Academy of Sciences |
| ZFMK |
Zoologisches Forschungsmuseum Alexander Koenig |
| NMNH |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Scelimeninae |
|
Tribe |
Discotettigini |
|
Genus |
|
|
SubGenus |
Discotettix |
Discotettix (Discotettix) belzebuth ( Serville, 1838 )
| Skejo, Josip, Pushkar, Taras I., Kasalo, Niko, Pavlović, Marko, Deranja, Maks, Adžić, Karmela, Tan, Ming Kai, Rebrina, Fran, Muhammad, Amira Aqilah, Abdullah, Nurul Ashikin, Japir, Razy, Chung, Arthur Y. C. & Tumbrinck, Josef 2022 |
Discotettix adenanii Mahmood et al., 2007: 1276
| Mahmood, K. & Idris, A. B. & Salmah, Y. 2007: 1276 |
Discotettix belzebuth: Bolívar 1887: 306
| Tan, M. K. & Wahab, R. A. 2018: 123 |
| Kuravova, K. & Wahab, R. A. & Kocarek, P. 2017: 120 |
| Kocarek, P. & Kuravova, K. & Musiolek, D. & Wahab, R. A. & Kahar, S. R. A. 2015: 289 |
| Otte, D. 1997: 32 |
| Yin, X. - C. & Shi, J. & Yin, Z. 1996: 866 |
| Blackith, R. E. 1992: 46 |
| Steinmann, H. 1970: 216 |
| Gunther, K. 1938: 301 |
| Willemse, C. J. M. 1930: 8 |
| Hancock, J. L. 1913: 39 |
| Kirby, W. F. 1910: 2 |
| Hancock, J. L. 1907: 6 |
| Hancock, J. L. 1907: 213 |
| Rehn, J. A. G. 1904: 670 |
| Bolivar, I. 1887: 306 |
Tettix belzebuth: Stål 1873: 152
| Stal, C. 1873: 152 |
Discotettix armatus
| Costa, A. 1864: 59 |
Tetrix belzebuth
| Serville, J. G. A. 1838: 759 |