
Cynodictis peignei, Verger & Solé & Ladevèze, 2020
|
publication ID |
https://doi.org/ 10.5252/geodiversitas2020v42a16 |
|
publication LSID |
urn:lsid:zoobank.org:pub:0DF5ED60-2119-4F79-9F7A-15D86A00F0B4 |
|
persistent identifier |
https://treatment.plazi.org/id/6246667C-B270-404F-A371-93349B7C7A18 |
|
taxon LSID |
lsid:zoobank.org:act:6246667C-B270-404F-A371-93349B7C7A18 |
|
treatment provided by |
Valdenar |
|
scientific name |
Cynodictis peignei |
| status |
sp. nov. |
Cynodictis peignei n. sp.
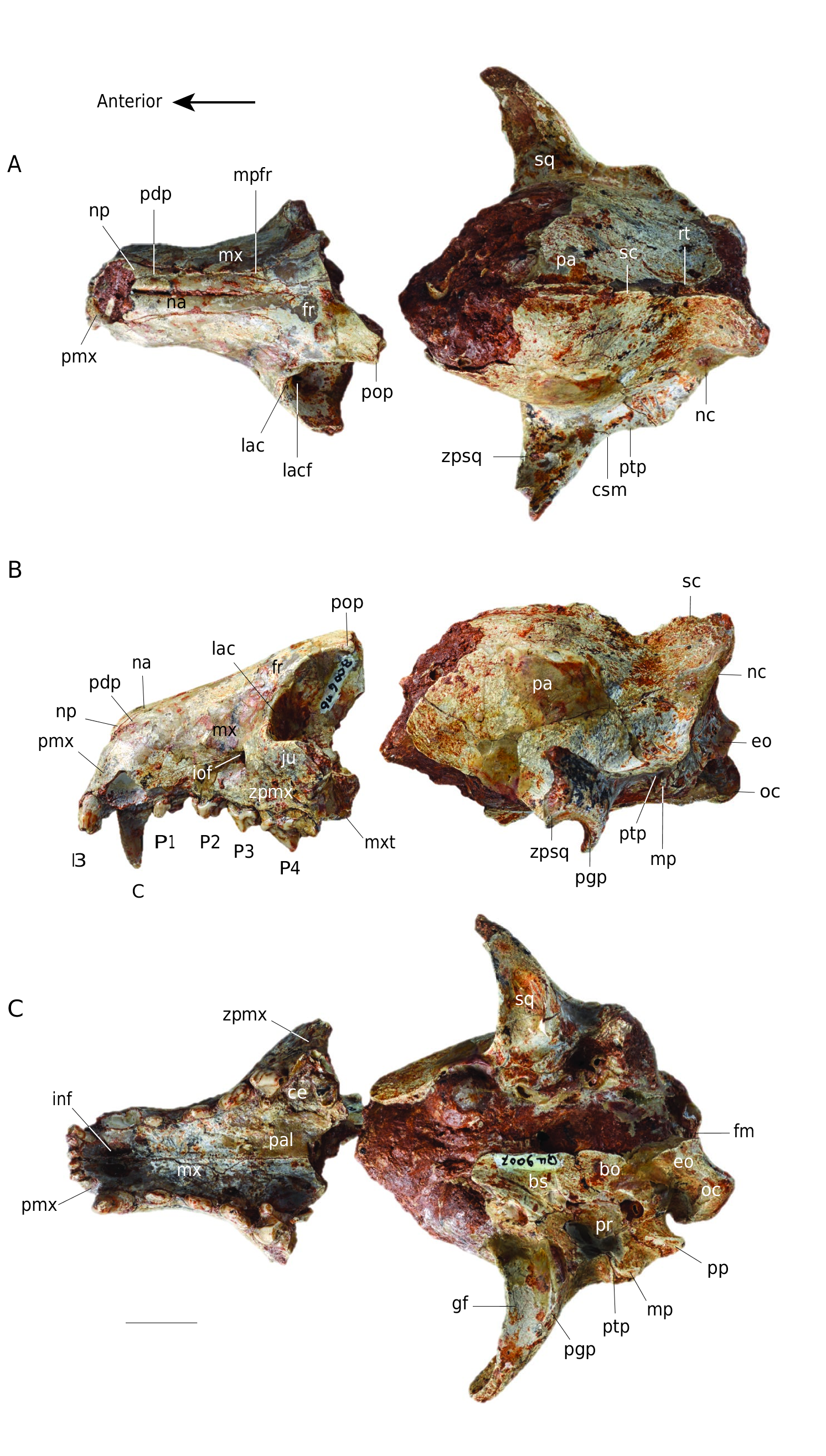
( Figs 2-4 View FIG View FIG View FIG )
urn:lsid:zoobank.org:act:6246667C-B270-404F-A371-93349B7C7A18
ETYMOLOGY. — Dedicated to the memory of our colleague Dr. S. Peigné (1972-2017), who described numerous carnivorous mammals from the Paleogene and Neogene of Eurasia and Africa.
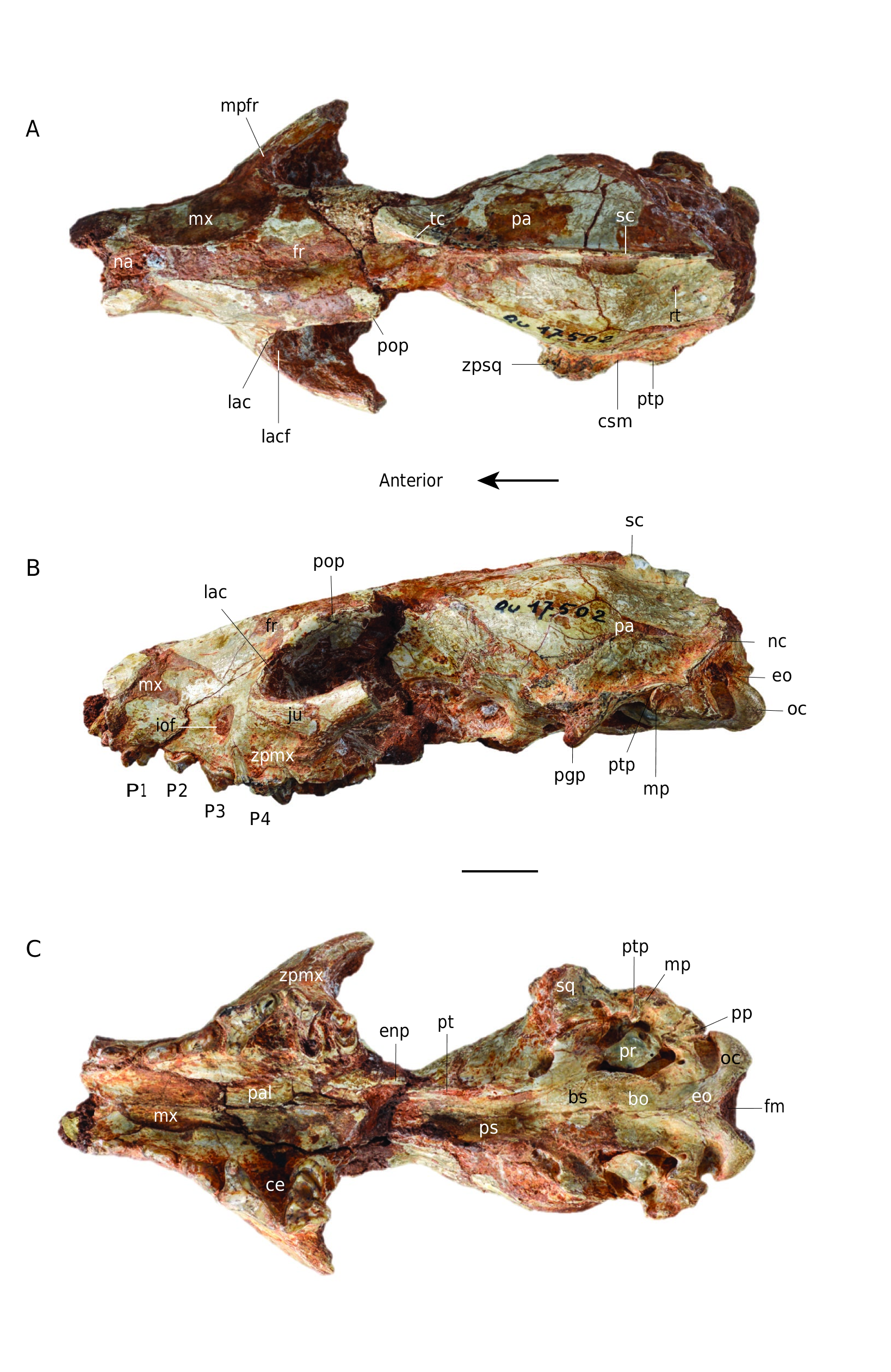
HOLOTYPE. — Incomplete cranium in two parts: MNHN.F.Qu9007 (snout) and MNHN.F.Qu9008 (neurocranium), with I1 -M1 left and I1 -P4 right (P4 broken).
TYPE LOCALITY AND HORIZON. — Quercy Phosphorites (old collection), estimated as late Eocene to early Oligocene (see Thenius [1959] and discussion of Kotsakis [1980: 268, 269]). It is noteworthy that the Mouillac deposit is no longer recognized since it is a mixture of phosphate bags, and the associated fauna gives no clue about the relative age of the specific Mouillac site.
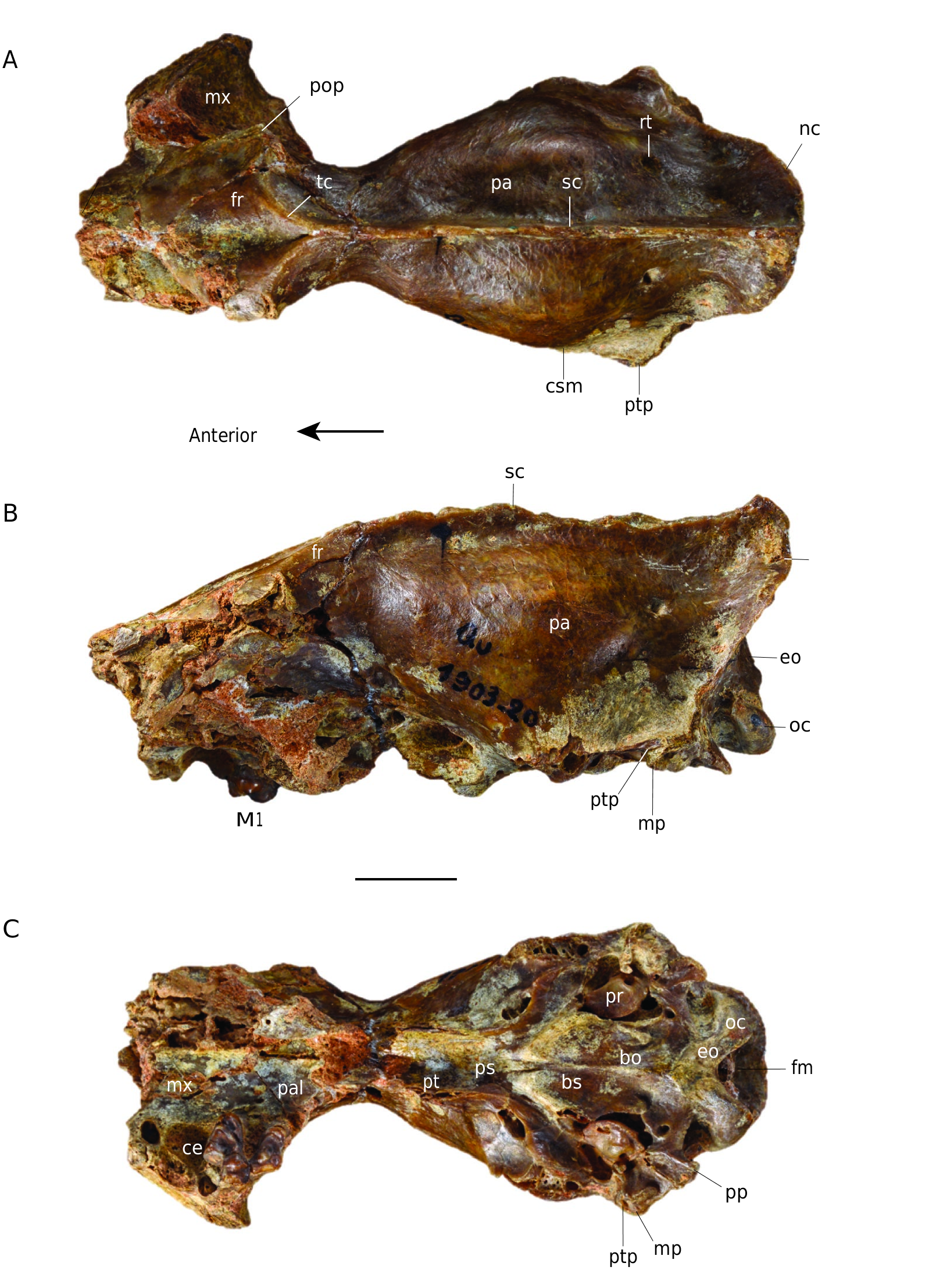
DIFFERENTIAL DIAGNOSIS. — The new species of C. peignei n. sp. is exclusively known by its cranium and its comparison to other Cynodictis species (diagnosed on the basis of lower teeth characters) is consequently limited to those known by cranial remains. Only two crania have been assigned to mandibular remains, belonging to two species: C. lacustris (MNHN.F.Qu17502) ( Teilhard de Chardin 1915: pl. II – former C. intermedius), and C. exilis (MNHN.F.Qu unnumbered).
C. peignei n. sp. cannot be compared to most of the existing other species of Cynodictis because they are not known by cranial remains: C. cayluxensis, C. ferox, C. crassus, C. longirostris. For the latter, Teilhard de Chardin (1915) illustrated in occlusal view a piece of a right maxilla bearing P2-M1, stored at the Montauban Museum, which he recognized as Cynodictis (ṙ) longirostris. Even though this determination is not certain, C. peignei n. sp. differs from this specimen by: a larger diastema between P2 and P3; P2 and P3 longer; P3 with a very strongly defined cingulum behind the accessory cusp; P4 very similar but with a smaller width at the base of the protocone; M1 with a straight stylar shelf on the labial edge of the tooth in occlusal view (central curvature on this same edge because of the development of the paracone and metacone in the Montauban Museum specimen); cingulum of M1 on the lingual edge forming a well-marked fossa between the latter and the protocone.
C. peignei n. sp. differs from C. exilis by: its much larger and stronger cranium; contact between the posterodorsal process of premaxillary and the lateral edges of the nasal more posterior (at the level of the canine in C. exilis); infra-orbital foramen at the level of P3 (posterior margin of P 3 in C. exilis); weaker transverse elongation of the zygo- matic arches; lacrimal foramen twice the size; post-tympanic process of squamosal less anteroventrally oriented; paroccipital processes proportionally taller and exoccipital wider; tensor tympani fossa larger; roof of the external acoustic meatus much deeper; nuchal crests almost vertical; braincase proportionally larger (despite the size difference); foramina for the ramus temporalis much smaller and closer to the sagittal crest; P3 much higher than P2 (P2 and P3 almost the same height in C. exilis); accessory cusp of P3 much larger; P4 with a narrower protocone area; metastyle of P4 shorter.
C. peignei n. sp. differs from C. lacustris by: a less transverse elongation of zygomatics arches; post-tympanic process of squamosal less vertically oriented; larger exoccipital despite a similar paroccipital process; larger tensor tympani fossa (although C. lacustris has a larger tensor tympani fossa than C. exilis); roof of the external acoustic meatus larger and deeper (it is the narrowest and shallowest in C. lacustris compared to C. peignei n. sp. and C. exilis); nuchal crests more vertically oriented (the nuchal crest of C. lacustris even hides the occipital condyles); much larger braincase; foramen for ramus temporalis closer to the sagittal crest (even closer in C. lacustris than in C. exilis); protocone area of P4 narrower; P4 metastyle shorter; M1 with a more rectangular shape; stylar shelf of M1 narrower, shorter and less posteriorly oriented; M1 metaconule more prominent; M1 protocone less prominent and M1 metacone higher than the paracone.
MEASUREMENTS. — See Table 2 and Table 3.
DESCRIPTION
Cynodictis peignei n. sp. is about twice as large as other species of the genus. Although broken at the level of postorbital constriction, the cranium is taller, wider and longer than other crania referred to Cynodictis . For instance, the braincase of the holotype is twice as large as that of C. lacustris. This large size recalls C. longirostris ( Filhol 1876; Teilhard de Chardin 1915; Bonis 1978), which is considered a large species of Cynodictis and notably characterized by the lengthening of the dentary. Cynodictis peignei n. sp. is, however, larger than this species. The specimen has lost most of the zygomatic arches, and parts of the neurocranium and of the inner wall of the orbit are missing. The right I1-P4 (incomplete P4) and the left I1-M1 (incomplete M1) are present. The skull has many fused bones. Very few sutures are clearly visible. The clearest sutures correspond to the following junctions: premaxillarymaxillary; premaxillary-nasal; maxillary-nasal; maxillarypalatine; maxillary-frontal; nasal-frontal; exoccipital-petrosal; squamosal-petrosal; basioccipital-basisphenoid. The lacrimalmaxillary and exoccipital-supraoccipital contacts can also be faintly distinguished. In addition, the teeth are all permanent and show significant wear. From all these observations, we consider this specimen to be an adult individual ( Thomé & Geiger 1997; Rager et al. 2013).
Dorsal view ( Fig. 2A View FIG )
The cranium of Cynodictis peignei n. sp. is composed of three parts: the snout, the orbito-temporal region and the braincase. The snout has parallel edges, which gives it a rectangular shape. The premaxillary is high and extends well beyond the anterior part of the nasal. Its posterodorsal process reaches as far caudal as the level of the P2. The maxillary extends onto the orbit. It shows slight lateral bulges in its anterior part, corresponding to the root of the canine. The infraorbital foramen, located at the level of the P3, is very wide and transversely extended. The nasal bones extend from the distal end of the snout (at the anterior border of the canine) to the frontal bones, where they are U-shaped. The frontal is not complete but seems large in C. peignei n. sp. A very small depression is visible on its midline. The maxillary process of the frontal is strongly developed anteriorly, reaching the anterior level of the P3. The post-orbital constriction is not preserved and cannot be described. The orbito-temporal region is very poorly preserved. The zygomatic arches, including the jugal and the zygomatic process of the squamosal, are not preserved. The base of the squamosal zygomatic process in the posterior portion of the temporal fossa is perpendicular to the anteroposterior axis of the skull. The zygomatic arch ends at the contact of the squamosal with the alisphenoid and parietal bones; its posterolateral end shows a marked supramastoid crista and is curved posteriorly. The supramastoid crista joins the post-tympanic process of the squamosal, the latter extending posteriorly to the nuchal crest. These crests have an oblique orientation with respect to the dorsoventral and anteroposterior axes. The sagittal crest is incomplete. It originates at the level of the connection between the two temporal ridges and joins the contact between the two nuchal crests corresponding to the occipital protuberance. The nuchal and sagittal crests are well developed in C. peignei n. sp. The bulge of the parietal reflects the size of the lyreshaped endocranium. On the parietal, close to the occipital protuberance, there is a well-marked foramen, on both sides of the sagittal crest, which corresponds to the passage of the ramus temporalis of the facial nerve.
Lateral view and internal wall of the orbit ( Fig. 2B View FIG )
In this view, the skull appears more elongated than high. The height of the skull increases only slightly from front to back. The maxillary is broad and slightly domed under the orbit. The lacrimal, preserved on the left side of the cranium, is in contact with the frontal, maxillary, palatine, and jugal. The lacrimal is a small bone with slightly visible wavy sutures, and which has a relatively large lacrimal foramen, filled on the left by sediment, but visible on the right side. More or less circular, it is located on the medial edge of the jugal above the maxillary foramen (the internal orifice of the infraorbital foramen). The maxillary foramen is about three times larger than the lacrimal foramen. The post-orbital process, which is located posterodorsally to the lacrimal and frontal and represents the posterodorsal limit of the orbit, forms a prominent point on the left side. The maxillary tuberosity is rather weak. The palatine, in its most anterior part (i.e., at the junction with the lacrimal and the maxillary), has only one foramen. The quality of preservation does not make it possible to know exactly whether it is the caudal palatine foramen or the sphenopalatine foramen (not illustrated here). The rest of the inner wall of the temporal fenestra is not preserved. On the posteroventral part of what is preserved of the zygomatic arch, the post-glenoid process is well marked. It is very slightly curved forward and thus forms the floor of the glenoid fossa. Posterior to the post-glenoid process and just posterior to the external acoustic meatus, the squamosal presents a very slightly developed post-tympanic process, which is very strongly anteroventrally oriented. It is joined by the mastoid process of the petrosal, which is half as small and points ventrally. Posterior to the mastoid process is the paroccipital process (jugular process of Evans 1993 = paracondylar process of the exoccipital of Wible & Spaulding 2013). This process is well developed and posteroventrally oriented. The exoccipital forms a ventral condyloid pit between the paroccipital process and the occipital condyle. The latter is rather broad and oriented in the same way as the paroccipital process. The two occipital condyles form the foramen magnum, which is wider than high. In lateral view, the connection between the nuchal and sagittal crests provides a very wide area for insertion of the temporal and nuchal muscles.
Ventral view ( Fig. 2C View FIG )
Laterally, the premaxillary ends in front of the canines, while its posterior extension forms a point ending posterior to the canines. The premaxillary has two incisive foramina and one interincisive foramen. The former have a teardrop shape, while the latter is much smaller. The palatine is partially damaged, but the maxillary-palatine contact is distinguishable, it starts at the anterior edge of the P4. The palatine, anteriorly rounded, forms a shelf delimited laterally by the P4 and M1. The major palatine foramina are no longer distinguishable. The posterior portion of the palatine, the presphenoid, the pterygoid and the anterior part of the basicranium are not preserved. Two foramina of great size, and in the same depression at the base of the alisphenoid, are visible laterally on the right side of the skull. The anterior-most foramen corresponds to the caudal opening of the alisphenoid canal. The posterior-most one corresponds to the foramen ovale. It is oriented obliquely and is opposite to the glenoid fossa. Laterally to these foramina, the squamosal bears the glenoid fossa, which is very elongated transversely. The condylar process of the mandible articulates in this pit. The basisphenoid and basioccipital are altered and barely distinguishable. The tubercle bordering them, where the longus capitis muscle attached, is not preserved.
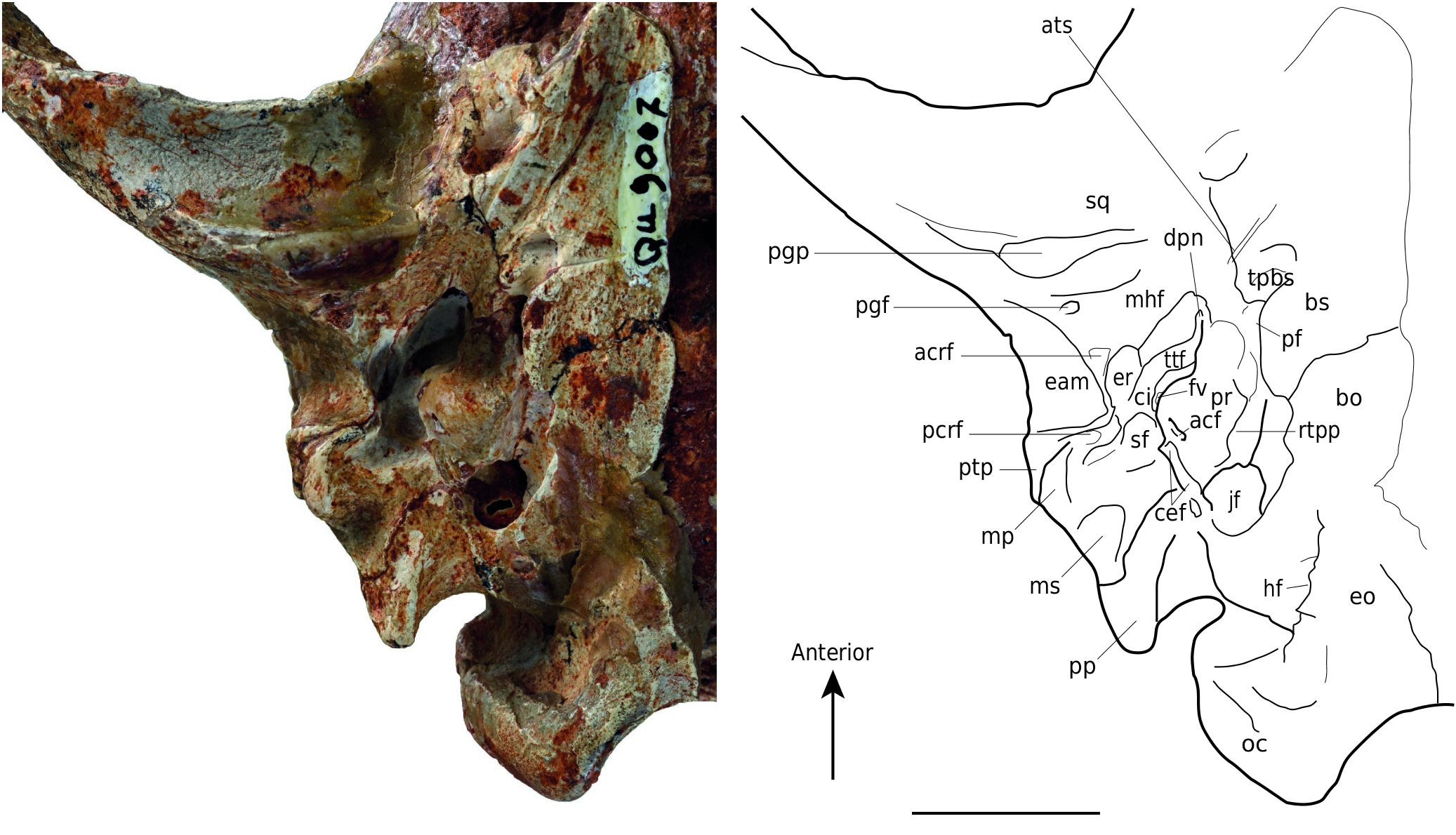
Auditory region ( Fig. 3 View FIG )
At the posterior part of the post-glenoid process and close to the lateral edge of the skull is the post-glenoid foramen of the squamosal. Posterior to it and posteromedially located in the tympanic cavity, a deep and narrow depression corresponds to the petrotympanic fissure (from which emerges the chorda tympani). Medially to the latter and bordering the (incomplete) tegmen tympani of the petrosal, a wide anteroposteriorly stretched depression is probably a facet for the insertion of the spine of the rostral process of the malleus (as described and illustrated in Nandinia Gray, 1830 by Wible & Spaulding 2013). Laterally to the petrotympanic fissure, a smaller but deeper depression, just posterior to the post-glenoid foramen, likely received the anterior crus of the ectotympanic, the external element of the auditory bulla. Near its external edge, a large and broad bony shelf formed by the squamosal corresponds to the roof of the external acoustic meatus. It is bordered anterolaterally by the post-tympanic process of the squamosal, on which there is a facet for the insertion of the posterior crus of the ectotympanic, which is attached posteriorly to the mastoid process of the petrosal. The petrosal is characterized in ventral view by an anterior bean-shaped part that is stretched anteriorly, the promontorium, and a posterior tongue-like part, the mastoid. The promontorium is slightly rough on its lateral and central surfaces. Its anterior extension is elongated and rounded. It has a transverse groove for the passage of the internal carotid artery.
The promontorium of C. peignei n. sp. is pierced at its posterior base by two foramina: the opening of the cochlear fossula, which contains the fenestra cochleae, and, anterodorsally to the lateral rim at the cochlear fossula, the vestibular fossula (fenestra vestibuli), more dorsal and rounded, which housed the footplate of the stapes. The opening of the cochlear fossula is directed towards the mastoid plate of the petrosal. The cochlear fossula is anteriorly overlapped by a bulge of the promontorium resulting from the first turn of the underlying cochlea (= tympanic ramp). The latter is connected to the tympanic cavity by the secondary tympanic membrane, housed in the cochlear fossula. The posterior extension of the cochlear fossula forms a broad depression stretched transversely and laterally bordered by a short process, interpreted here as the medial section of the caudal tympanic process (sensu MacPhee 1981). The vestibular fossula, which connects the ossicular chain to the vestibular ramp of the cochlea, is located anterior to the probable level of the tympanohyal (not preserved here) and opens towards the roof of the external acoustic meatus. These two openings are separated by the crista interfenestralis. The mastoid part of the petrosal is delimited anterolaterally by the mastoid process, which forms a narrow transversely and ventrodorsally elongated shelf. Posterior to the mastoid process, a bean-shaped osseus plate is delimited medially by a broad shelf of the mastoid that is very slightly concave, almost flat, and smooth. This plate corresponds to the mastoid exposure. The shelf continues medially to the exoccipital and participates in the prominent paroccipital process of the exoccipital.
Posteroventrally, the paroccipital process of C. peignei n. sp. is hollowed out on its anterolaterally oriented inner face. It bears two ridges: the most mesial extends from the apex of the process to the cochlear fossula; the most lateral (more altered) extends from the same apex to the mastoid process, with a slight curve. The medial margin of the mastoid shelf forms a thin bony process surmounted by a bulge in front of the cochlear fossula. It corresponds to the lateral section of the caudal tympanic process (sensu MacPhee 1981). If the tympanohyal was present it would form, with the caudal tympanic process, a very rounded notch, the stylomastoid foramen (not preserved here). More dorsally, the stapedius fossa is deep, more or less oval and its anterior wall is formed by the gyrus (a cerebral convolution) of the underlying semicircular lateral canal. It indicates the location of the stapes and stapedial muscle. Anterolaterally and medially delimited by the promontorium, is a wide and deep depression, slightly deteriorated. The bony roof of this depression consists partly of an epitympanic wing of the petrosal (particularly altered here) anteriorly, and an epitympanic wing of the squamosal and tegmen tympani posteriorly. The anterior-most cavity, which is oriented transversely, is the pit for the tensor tympani muscle, attached to the tympanic membrane and whose function is to dampen the sounds and houses the “nape” of the malleus. The tensor tympani fossa is separated from a more posterior and rounded depression by a small bony wall. This depression is hollowed out by two fossae. The most anterior one, which is also the widest, is the epitympanic recess. Essential in the proper functioning of the ossicular chain, the epitympanic recess housed the malleus-incus articulation. The most posterior pit, the fossa incudis, is half the size but deeper than the epitympanic recess. It housed the short process of the incus. It is located anteriorly to the stapedius fossa and is separated from it by the crista parotica, which forms a thick bone barrier.
Medial to the epitympanic recess and the fossa incudis is the damaged facial canal. This canal runs along the medial edge of the promontorium. It opens between the epitympanic recess and the vestibular fossula. On the medial edge of the promontorium and on the most medial part of the posterior bulge of the cochlear fossula are two very distinct facets that receive the caudal entotympanic (an element of the auditory bulla). The most anterior and longest facet is on the rostral tympanic process of the petrosal. The bony margin marking the posterior border of the cochlear fossula is attached to two marked tubercles of the exoccipital. These two tubercles delimit two grooves, of which the most posterior probably marks the passage of the vagus nerve (X).
Anteromedially to these two tubercles there is a large foramen corresponding to the jugular foramen. The jugular foramen is included in a long fissure – enlarged in the specimen because of a taphonomic deformation –, which extends between the promontory and the basioccipital. The hypoglossal foramen pierces the exoccipital, and is located posteromedially in the jugular foramen. The promontorium apex is medially separated from the basioccipital by a very large hole, which may correspond either to the piriform fenestra (sensu MacPhee 1981), or to the foramen lacerum of Evans (1993) (see the discussion concerning this structure in Wible & Spaulding 2013). This orifice contains the foramen for the internal carotid artery. At the front of this large hole, the tympanic process of the basisphenoid forms a large bone pocket. The basioccipital is too altered to observe an excavation as in other Cynodictis skulls (KLV pers. obs.). Located between the tympanic process of the basisphenoid and the sulcus for the auditory bulla, a wellmarked groove begins at the level of a foramen just anterior to the apex of the promontorium. This foramen probably marks the passage of the deep petrosal nerve.
Anterior
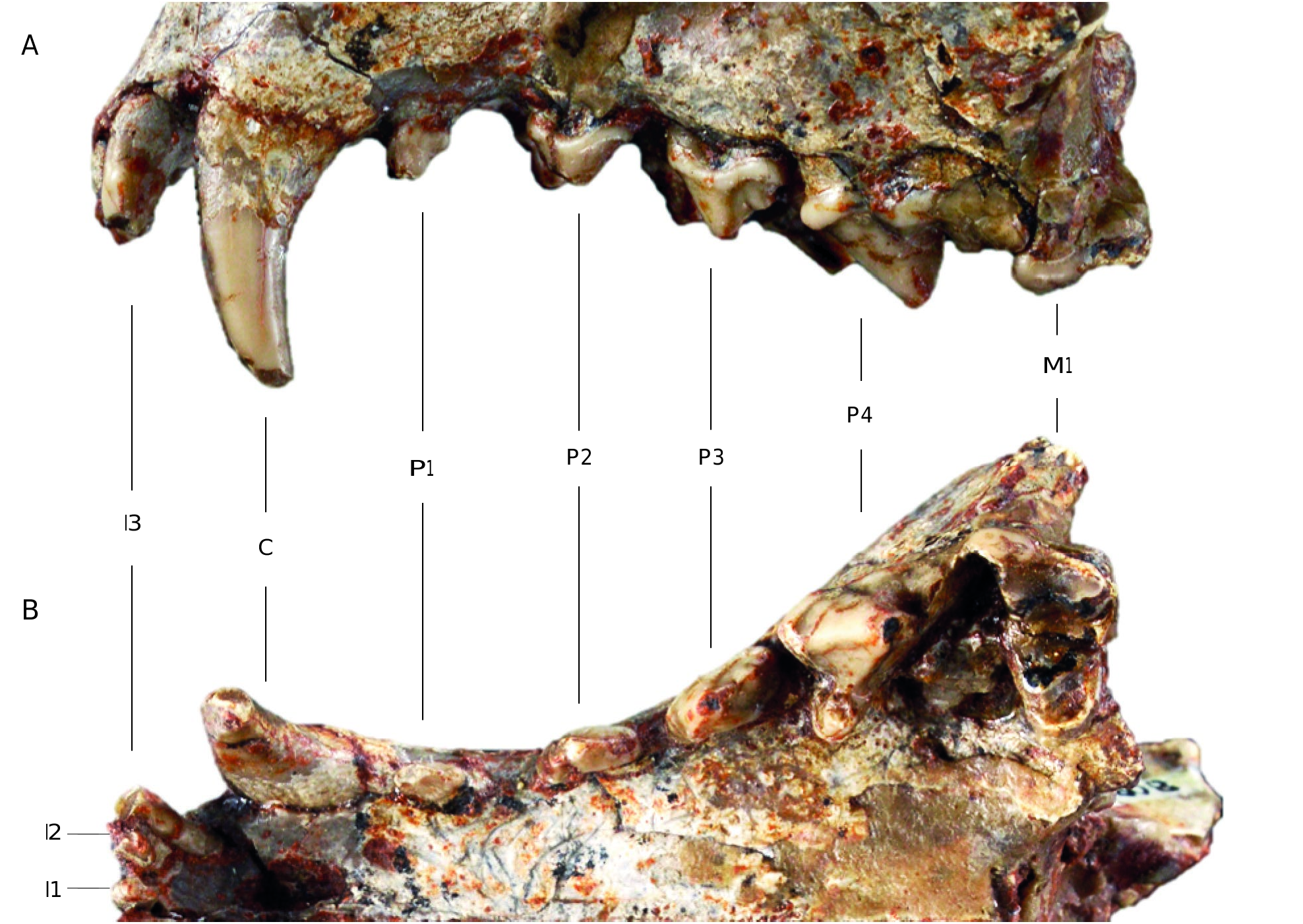
Upper teeth ( Fig. 4 View FIG )
The specimen described here was found with two hemimandibles (MNHN.F.Qu9009 and MNHN.F.Qu9010). However, the lower and upper teeth do not occlude properly. This implies that they do not belong to the same specimen ( Crompton & Hiiemäe 1969). The right I3, M1, and M2, and the left M2 are not preserved. Moreover, the left M1 and the right P4 are badly damaged. The I1 and I2, separated by a very slight diastema, are smaller than the I3, which is twice as large. The three incisors are conical, rectangular and single-rooted. A very small diastema separates them from the canines. The canines are conical and very slightly curved towards the back. A diastema of about the same length as the one that separates the incisors from the canines is present between the canine and the P1. The latter is single-rooted and has a posterior accessory cusp. The largest diastema separates the P1 from the P2. In lateral view, the teeth are two-rooted, taller, and longer from P2 to P4. The P2 is conical and has a single prominent cusp, the paracone. Its cingulum is very thin, but almost complete. It has a very weak cusp mesially and a stronger one distally. The P3, higher than the P2, has the same morphology as the P2 but differs by having acces- sory cusps that are more developed and individualized. The first one, which is rounded, is located posterior to the main cusp (= paracone). This accessory cusp and the paracone are connected by a short but well-developed ridge. The second accessory cusp is much smaller and is located anterior to the main cusp. The P4, whose anterior root forms a bulge on the maxillary, has a large oblique cingulum at its anterior base. The paracone is, by far, the tallest cusp of all the premolars. It points backwards and exhibit a posterior ridge as well as an anterior crest. The posterior crest reaches the (incomplete) metastyle, which is long, protruding, and shows a very slight concave curvature at its center. Its contact with the paracone is lingual relative to the middle of the tooth, orienting the metastyle towards the posterior part of the skull. The carnassial notch is present between the paracone and metastyle. The P4 has a fairly large lingual shelf, which carries a welldeveloped protocone. The M1 is rectangular in shape and partially worn. The cingulum is well developed on the lingual part of the talon, where it forms a very strong bulge that is narrow mesiodistally. The stylar shelf, much smaller than the protocone, is oblique orientated outward with respect to the anteroposterior axis. The stylar shelf includes three cusps. The
A
metastyle is poorly developed, unlike the parastyle that forms a prominent and strongly rounded cusp. Both the metacone and paracone are prominent and sharp, the metacone being the tallest. The centrocrista and the paracrista are more salient than the metacrista. The talon is very broad and points towards the buccal part of the oral cavity (but also with a slight posterior inclination). The protocone is very strong, eroded and is mesially shifted. Close to the protocone, there is a metaconule, but no protoconule is visible (absent or worn down). The preprotocrista is more marked than the postprotocrista and reaches the parastyle. Two very thin cingulae are visible on the lingual base of the metacone and the paracone.
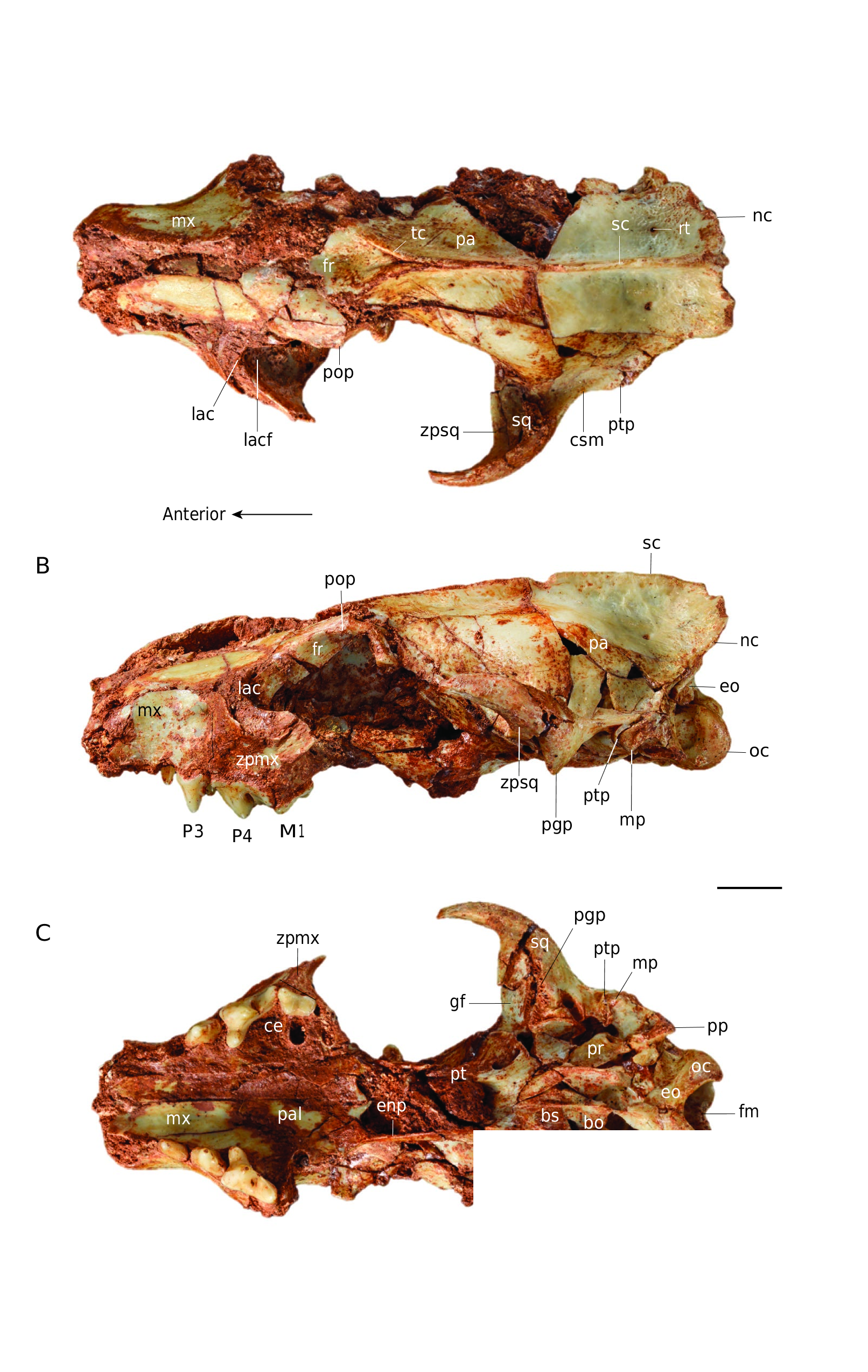
Comparison ( Figs 5-7 View FIG View FIG View FIG )
The posterodorsal process of the premaxillary of C. peignei n. sp. contacts the lateral edges of the nasal further back than in other Cynodictis species. The premaxillary ends laterally at the posterior level of the canine, whereas it ends at the anterior third of the canine in the other Cynodictis species. The infra-orbital foramen of other Cynodictis species is located at the anterior edge of the P4. In C. lacustris, the posterior part of the nasal bones is V-shaped rather than U-shape as seen in C. peignei n. sp. The maxillary process of the frontal also differentiates these two species: its tip stops at the infra-orbital foramen in C. lacustris. The snout of C. lacustris is more tapered than in the other species. In C. lacustris, the face abruptly increases in transverse width. This transverse elongation occurs at the level of the infra-orbital foramen and is due to the separation of the zygomatic processes. This transverse elongation is weaker in C. peignei n. sp. The snout of C. peignei n. sp. is higher than
nc
in C. lacustris, but, surprisingly, it is not longer. The lacrimal foramen of C. peignei n. sp. is larger than in other Cynodictis species. The base of the zygomatic process of the squamosal (in the posterior portion of the temporal fossa) forms a forward curvature in all Cynodictis species. In C. lacustris and C. exilis, the post-glenoid process is more forwardly curved; C. lacustris has the greatest curvature, pointing almost 30° anteroventrally. In C. exilis, the supramastoid crista is flatter, more pronounced and concave, than in C. peignei n. sp. The post-tympanic process of the squamosal is more vertically oriented in C. peignei n. sp. than in C. lacustris and C. exilis. The mastoid process is more rounded and points more laterally. The paroccipital processes of C. peignei n. sp. are shorter and the exoccipital is thicker than in other Cynodictis species. The promontorium is relatively similar in size but is more massive and less triangular than in C. exilis and C. lacustris. In C. lacutris and C. peignei n. sp., the hypoglossal foramen is closer to the petrosal in comparison to the other Cynodictis species. The tensor tympani fossa of C. peignei n. sp. is larger than in the other Cynodictis species. On the other hand, the insertion of the anterior crus of the ectotympanic is relatively smaller in C. peignei n. sp. The rostral tympanic process of the petrosal forms a protuberance in C. peignei n. sp. The roof of the external acoustic meatus is much deeper in C. peignei n. sp. than in other Cynodictis species. A major difference should be emphasized: in C. peignei n. sp., the nuchal crests are almost vertical and do not mask the condyles, whereas all other Cynodictis species, the nuchal crests are very strongly elongated posteriorly to the point at which they completely hide the occipital condyles in dorsal view. The braincase is much larger in C. peignei n. sp. The foramina for the ramus temporalis are smaller and closer to the sagittal crest than in other Cynodictis species. The condyles are more prominent in C. peignei n. sp. than in C. exilis. In C. peignei n. sp., they are twice as large, resulting in a deeper ventral condyloid fossa. The tubercle lying in the basioccipital and basisphenoid is never complete in our sample, but it should be noted that this structure in C. lacustris is much more strongly developed than in C. peignei n. sp. and C. exilis. There is no particular difference between the P1, P2 and P3 of the Cynodictis specimens of the sample, except the presence only in C. peignei n. sp. of a P3 higher than the P2, and of a more developed accessory cusp on P3. The P4 has a narrower protocone area, more anteriorly oriented than in other Cynodictis species. The metastyle of the P4 of C. peignei n. sp. is shorter than in the other species. The M1 of Cynodictis peignei n. sp. differs from the other Cynodictis species by its less trapezoidal and more rectangular shape. A connection between the stylar shelf and protocone area is absent in the studied specimen. Its stylar shelf is narrower, shorter, and oriented less posteriorly. The metaconule is more prominent, while the protocone is less prominent than in the other Cynodictis species. The metacone is taller than the paracone in C. peignei n. sp. which is not the case in other species.
RELEVANCE OF CRANIAL CHARACTERS
Among the extinct mammal species described and named so far, a very large number have been defined only based on dental characters. This is illustrated by the diagnoses proposed for almost all mammalian groups (except for Xenarthra and Pholidota because of their reduction of teeth). This is due to the nature of the fossil record: dental elements are abundant because they have been preserved and fossilized due to the mineralization of their tissues. Moreover, dental elements can provide information on the diet, as well as on the body mass of extinct species.
Amphicyonidae are clearly not an exception (see the diagnoses proposed by Springhorn 1977). The characters considered as diagnostic are substantially concentrated on the tooth row and more specifically on the morphology of the m1. However, this type of character has a considerable number of biases because molars have at least three constraints for identification and descriptive studies:
1) Although dental structures are a relevant ontogenetic index – even if it is questionable for some groups ( Ciancio et al. 2011) –, the patterns of wear and the phenotypic plasticity of the teeth result in morphological variability and may cause problems with fossil species discrimination ( Gingerich 1974; Suchentrunk & Flux 1996; Tsoukala 1996; Hillson 2005).
2) Functional constraints on the teeth result in a large number of convergences. This case is well illustrated with the debate on the position of Amphicyonidae . The “bear-dogs”, so named because of their anatomy (dentition and locomotion), which is sometimes similar to that of Canidae , sometimes to that of Ursidae , represent a phylogenetic enigma ( Viranta 1996). Historically, they were first considered to be very close to Canidae ( Matthew & Granger 1924; Petter 1966), a hypothesis that has been reconsidered recently ( Spaulding & Flynn 2012). They have also been considered as the sister group of Ursidae based on many morphological characters ( Ginsburg 1966; Hough 1948; Hunt 1977; Wyss & Flynn 1993) or as the sister group of Arctoidea (the group that gathers ursids, pinnipeds, and musteloids; Finarelli 2008; Hunt 1996, 1998). Finally, the most recent studies regard the “bear-dogs” as the sister-group of all Caniformia ( Tomiya & Tseng 2016; Wesley-Hunt & Flynn 2005). Other examples are present in the history of the caniforms. For example, the case of Musteloidea, where the whole group presents a wide range of dentition and locomotion, resulting in difficulties for the paleontologists to discriminate the different groups ( Law et al. 2018).
3) The teeth are subject to serial homology (the similarity of repeated structures within an organism). This calls into question the characterization of a group based on repeated structures. For example, if a group is characterized by the presence of a cingulum on the M1, M2, and M3, then because all these characters are dependent on each other, they should be considered as representing only one characteristic that defines the group, and not as three independent characteristics ( Billet & Bardin 2018).
Although these problems seem alarming, we do not aim to question here previous studies because they are based on dental structures: indeed, several dental features used in these publications are not affected by these constraints, and thus are diagnostic (e.g., highly specialized dentition). The goal of the present discussion is to question specific diagnoses based on very weakly defined dental variants, especially when the sampling does not allow study of intraspecific variation.
During more than 150 years, the diversity of the genus varied from six to nearly thirty species ( Filhol 1876; Schlosser 1902; Teilhard de Chardin 1915; Bonis 1978; Kotsakis 1980). With the description of C. peignei n. sp. herein, seven species are now recognized. The previous overestimation of the taxonomic diversity can be explained by the effect of the constraining dental particularities listed above on the variation within the group.
It is worth remembering that the lower teeth can be morphologically related to the upper teeth in an individual because of the functional links caused by the occlusion ( Crompton & Hiiemäe 1969). Dayan et al. (2002) showed that dental features within carnivoran populations are more variable than cranial traits. In addition, dental traits are strongly correlated with each other, just as cranial traits are correlated with each other, but these two sets are not highly correlated with one another ( Dayan et al. 2002). This implies that teeth and skull may be subject to different selective pressures and constraints (e.g., genetic, development, function) and, therefore, the study of these structures separately might allow for envisaging different scenarios. To conclude, and in ideal cases where most of the skeleton is available for a fossil specimen, paleontologists should try to define in a more comprehensive way the new species they are erecting and should propose detailed and compared diagnoses based on all parts of the skeleton and not teeth (or fragments of teeth) only.
| MNHN |
France, Paris, Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |