
Cryptosepalum korupense Burgt, 2014
|
publication ID |
https://doi.org/ 10.5252/a2014n1a7 |
|
DOI |
https://doi.org/10.5281/zenodo.5902367 |
|
persistent identifier |
https://treatment.plazi.org/id/3F618792-FFAE-B91F-FF0E-FA0953B7F95A |
|
treatment provided by |
Carolina |
|
scientific name |
Cryptosepalum korupense Burgt |
| status |
sp. nov. |
Cryptosepalum korupense Burgt View in CoL , sp. nov.
( Figs 1-4 View FIG View FIG View FIG View FIG )
Cryptosepalum korupense , sp. nov. is related to Cryptosepalum staudtii Harms. Cryptosepalum korupense , sp. nov. has (10-)14-21(-26) pairs of leaflets; C. staudtii has 10-14(-16) pairs of leaflets. The leaflets of C. korupense , sp. nov. are divided by the midvein into two unequal parts; near the apex of the leaflet, the proximal part (the part closest to the twig) is 3-4 × wider than the distal part (the part furthest from the twig). In C. staudtii the proximal part of the leaflet near the apex is only 1-1.5 × wider than the distal part. The flowers of both species are more or less similar. The fruits of C. korupense , sp. nov. are 10-18 cm long; the fruits of C. staudtii are 6-10 cm long.
TYPUS. — Cameroon. Southwest Region, south of Mundemba, forest near village Besingi , about 1 km downstream of bridge over Idu river, on the south bank, 4°55’16’’N, 8°54’04’’E, alt. 50 m, young fruits, 12.III.2007, van der Burgt & Motoh 922 (holo-, K [ K000460356 ] ; iso-, BR, G, MO, P, SCA, WAG, YA).
PARATYPI. — Cameroon. Southwest Region, Korup National Park , south of P transect, 4°59’N, 8°49’E, alt. 100 m, sterile, 28.III.2004, van der Burgt & Eyakwe 681 (WAG, YA) ; same loc., 5°0’N, 8°48’E, alt. 100 m, seedlings, 25.V.2007, van der Burgt & Motoh 941 ( BR, G, K, MO, P, SCA, WAG, YA) ; Korup National Park, P transect to Isangele Road , 4°58’N, 8°50’E, alt. 100 m, fr.& seedlings, 21.II.2008, van der Burgt, Pearce , Poundje & Yombo 1126 (K, MO, P, WAG, YA) ; Near Besingi village, about 1 km downstream of bridge over Idu river, 4°55’16’’N, 8°54’04’’E, alt. 50 m, sterile, 14.IX.2006, van der Burgt & Motoh 848 (K, MO, P, WAG, YA) ; same loc., fr., 20.V.2007, van der Burgt & Motoh 931 (K, MO, WAG, YA) ; same loc., fr., 29.IV.2007, Motoh 1 (BR, K, MO, P, WAG, YA) ; same loc., seeds, 30.VII.2007, Motoh 3 (K, YA) ; near Besingi village, about 1 km upstream of bridge over Idu river, 4°55’20’’N, 8°54’37’’E, alt. 50 m, 2.II.2010, Motoh 6 (K, YA) ; Mount Cameroon, Mokoko forest above Bonja village, 4°28’N, 9°06’E, alt. 100 m, sterile, 24.III.1993, Tchouto ( Mbatchou ) 621 (K, SCA) ; Mount Cameroon, Onge River , 4°18’N, 9°01’E, alt. 200 m, sterile, 10.X.1993, Tchouto ( Mbatchou ) 773 (K, SCA) ; Mount Cameroon, Mokoko , 4°27’N, 9°04’E, alt. 300 m, sterile, 22.V.1994, Thomas 10049 (SCA).
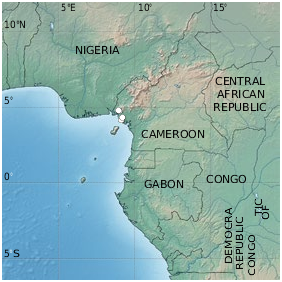
DISTRIBUTION. — Cryptosepalum korupense , sp. nov. is endemic in the Southwest Region in Cameroon ( Fig. 3 View FIG ). The species was found in southern Korup National Park, in the forest on the banks of the Idu River near the village Besingi, and in the lowland forest northwest of Mount Cameroon.
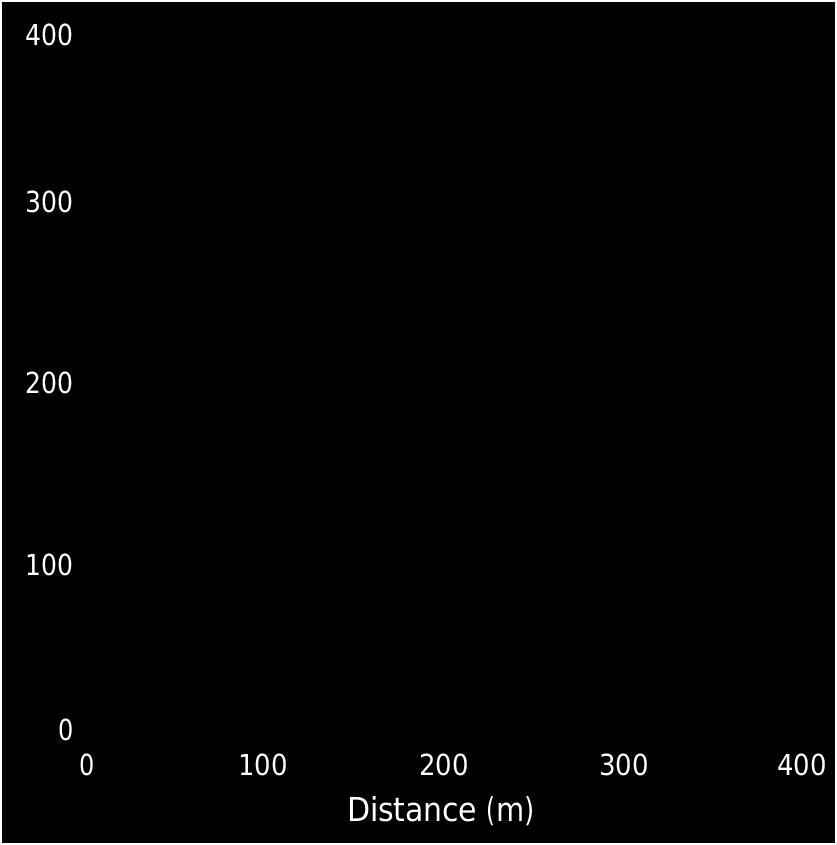
Three large plots have been made in southern Korup National Park; but C. korupense , sp. nov. was not found in any of these plots. These three plots are the plots “P”, 82.5 ha and “NW”, 56.25 ha ( Newbery et al. 2013); and the plot “KFDP”, 50 ha ( Kenfack et al. 2006). The new species seems to be rare in southern Korup National Park ( van der Burgt & Eyakwe 2010); which can also be concluded from the mapping of Microberlinia bisulcata trees. Three large groves of M. bisulcata have been mapped (one grove only partly); the area of forest mapped in and close to these groves is c. 7.9 km 2 (Newbery & van der Burgt unpubl. data). Within this area no C. korupense , sp. nov. trees were found. In addition, c. 66 km of parallel transect lines at 500 m distance to each other were walked along in southern Korup (Newbery & van der Burgt, unpubl. data). Any mature C. korupense , sp. nov. tree standing at less than 25 m from these transects would likely have been found by way of the usually abundant seedlings at this distance. This amounts to another 3.3 km 2 of forest mapped. Within this area three small groves were found; one group of 14 C. korupense , sp. nov. trees over 10 cm trunk diameter ( Fig. 4 View FIG ) and two small groups of two trees each. The total amount of forest mapped is c. 11.2 km 2; within this area 18 C. korupense , sp. nov. trees over 10 cm trunk diameter were found. Of all trees over 50 cm trunk diameter in southern Korup National Park, one tree out of c. 3100 trees is C. korupense , sp. nov. However, because these surveys were based on seedlings, mature trees which produced few or no seedlings in the years before these surveys, as well as immature trees, may have been overlooked.
In the forests near Besingi village ten Cryptosepalum korupense , sp. nov. trees with trunk diameters of 20-60 cm were recorded. All trees were found on the banks of the Idu River. This river is bordered by an oil palm plantation on one side and by a mosaic of subsistence farms and secondary forest on the other side. Along the river a strip of forest rich in rare forest species remains. This forest strip is a remnant of a once more continuous forest and has not been converted to farmland because of the often steep and rocky banks and variable water level.
HABITAT. — Rainforest on well-drained soil; 50-300 m.
ETYMOLOGY. — Named for Korup National Park where the species was first found.
SEED DISPERSAL. — Trees of Cryptosepalum korupense , sp. nov. appear to grow in small groups. The only group that could be mapped completely had 14 trees over 10 cm trunk diameter ( Fig. 4 View FIG ). Several other species of Caesalpinioideae are present within this group, as well as many trees from other families. The seeds of C. korupense , sp. nov. are dispersed by way of ballistic seed dispersal. The maximum ballistic dispersal distance is unknown but it is probably in the range of 50-70 m. This estimate is based on van der Burgt (1997) and on later observations by the first author on other Caesalpinioideae taxa with similar pod and seed size and thickness, and therefore presumably similar maximum dispersal distance. The tendency of trees of this species to grow in groups ( Fig. 4 View FIG ) is probably related to the relatively short and strictly limited maximum dispersal distance of the ballistic seed dispersal method.
CONSERVATION STATUS
Cryptosepalum korupense , sp. nov. is assessed here as Endangered, B1ab(iii;v) C1 D, under the criteria of IUCN (2013). The extent of occurrence of C. korupense , sp. nov. is 802 km 2. The forests in the area between the known localities are severely fragmented. Outside Korup National Park, which is a protected area, there is continuing decline in quality of habitat and in the number of mature individuals, due to subsistence farming and the establishment of oil palm plantations. These facts lead to Endangered under IUCN criterion B1 ( IUCN 2013).
In total 28 trees have been recorded in two of the three localities (one locality, the lowland forests northwest of Mount Cameroon, was not visited but taken from herbarium sheets). Assuming that trees become mature at 30 cm trunk diameter, 20 of the 28 trees are mature. Much forest in the Southwest region remains unexplored for this species; therefore the number of mature trees is expected to be more than 50. However, in southern Korup National Park the species is very rare (see the paragraph on distribution); therefore the number of mature trees may be less than 250, is leading also to Endangered, under IUCN criterion D.
The estimated number of mature trees, combined with the estimated decline of the species of at least 20% over two generations (which may be over 100 years), due to subsistence farming and the establishment of oil palm plantations, is leading also to Endangered, under IUCN criterion C1.
DESCRIPTION
Tree to 44 m high, trunk to 88 cm diameter at 1.3 m high, bole cylindrical to somewhat irregular in large trees, bark light grey-brown, smooth.Twigs densely tomentose to glabrescent, hairs curly, 0.3-0.7 mm long. Bud scales 14-18, caducous, distichous, not keeled, parallel-veined; proximal scale 1 mm long × 3 mm wide, distal scales progressively larger and relatively longer, to 40 × 18 mm, oblanceolate, bright dark pink; both surfaces sparsely hairy with hairs to 0.1 mm, margins with hairs to 1 mm. Stipules in pairs, free, caducous, parallel-veined, longpetioloid, oblanceolate, 20-40 × 1-10 mm; apex acute; indumentum similar to bud scales. Leaves alternate, paripinnate, (5.5-)7-11(-12) × (2-) 2.5- 3.2 cm; petiole 3-5 mm, without glands; rachis (4-)5.5-9.5(-11) cm long, densely tomentose, hairs curly, 0.3-0.7 mm long; upper and middle leaflet pairs opposite, lower pairs subopposite. Leaflets sessile, in (10-)14-21(-26) pairs; narrowly oblong, to 13-15 × 4-5 mm, base and apex asymmetric, apex emarginate; midvein dividing the leaflet into two unequal parts; at the base of the leaflet the proximal part (the part closest to the twig) is as wide as the distal part (the part furthest from the twig); near the apex of the leaflet the proximal part is 3-4 × wider than the distal part; leaflets glabrous both sides, caducous fringing hairs to 0.5 mm long, a few persistent hairs to 0.2 mm long on apex of midvein; glands 0-2, one gland positioned on distal part of leaflet, near the base and about midway between the midvein and the margin, on lowest leaflets also a gland on proximal part of leaflet. Inflorescence a raceme, axillary, one per node, 20-25-flowered, 5-7.5 cm long including a peduncle c. 2 cm long; densely tomentose, hairs curly, 0.3-0.7 mm long.
Flowers
Floral bract inserted at base of pedicel, caducous, lanceolate, c. 8 × 3 mm, indumentum similar to bud scales; pedicel 10-14 mm long, indumentum similar to inflorescence axis; bracteoles inserted at apex of pedicel, white, opposite, ovate, valvate, 6 × 5 mm, keeled, apex acute, mostly glabrous outside, short hairs outside near apex, on edge and inside; hypanthium campanulate, c. 0.8 mm deep, glabrous. Sepals 4, white, lanceolate; adaxial sepal 1-2 × 0.3-0.6 mm, sometimes bilobed, margins hairy; lateral sepals 0.2-1 × 0.3-0.4 mm; abaxial sepal 0.5-2 × 0.3-0.5mm. Adaxial petal white, glabrous, 6-7 mm long, 8 mm wide, claw 1 mm long; other petals absent. Stamens 3, filaments 8-9 mm long, glabrous, anthers oblong-elliptic, c. 2.2 × 1.5 mm. Staminodes 2-6, to 3 mm long. Ovary oblong, stipitate, 4.5 × 1.5 mm, glabrous, ovules 2-4; stipe 1-1.5 mm long, curled hairs to 1.5 mm long, placed on the sides; style 6-7 mm long, glabrous, stigma capitate. Pod oblong to obovate, glabrous, lacking surface sculpture; broadest towards apex, 10-18 × 3.5-5.5 × 0.3-0.8 cm, beak 3-8 mm long; upper suture broadened, to 3 mm wide per valve; containing 0-4 seeds. Seeds discoid, 22-28 × 16-22 × 4-5 mm. Seedling: hypocotyl 7.5-9.5 cm, epicotyl 2-3.5 cm, first pair of leaves opposite, leaves (6-)7.5- 10.5 × 2.5-3.5 cm, leaf rachis 6-10 cm, leaflets in (14-)16-22 pairs, to 18 × 6 mm.
IDENTIFICATION NOTES
The small, emarginate leaflets of Cryptosepalum korupense , sp. nov. have a unique character distinguishing them from all other West and Central African Caesalpinioideae species. The midvein divides the leaflet into two unequal parts; near the apex of the leaflet, the proximal part (the part closest to the twig; see Wieringa 1999: 40) is 3-4 × wider than the distal part (the part furthest from the twig). In other African Caesalpinioideae species with small, emarginate leaflets the proximal part of the leaflet near the apex is only 1-2 × wider than the distal part. This character can be used to distinguish C. korupense , sp. nov. from other species of Caesalpinioideae with small, emarginate leaflets; for example C. staudtii , Didelotia brevipaniculata J.Léonard , Microberlinia bisulcata and Plagiosiphon emarginatus . Flowering specimens of these species are easy to distinguish from C. korupense , sp. nov. (see Aubréville 1970).
Cryptosepalum korupense , sp. nov. can be easily distinguished from the other tree species of Cryptosepalum , because the leaves have more pairs of leaflets than any of the other tree species: (10-)14-21(-26) pairs. Cryptosepalum staudtii has 10-14(-16) pairs of leaflets; the other tree species are unifoliolate, bifoliolate or tetrafoliolate.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |