
Crellomima sigmatifera Fernandez, Gastaldi & Hajdu, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5052.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:FED635BA-B982-400E-B920-1DBA22025EA9 |
|
DOI |
https://doi.org/10.5281/zenodo.5572197 |
|
persistent identifier |
https://treatment.plazi.org/id/15003760-FF9B-F66A-6591-FB94FEDEFD1E |
|
treatment provided by |
Plazi |
|
scientific name |
Crellomima sigmatifera Fernandez, Gastaldi & Hajdu |
| status |
sp. nov. |
Crellomima sigmatifera Fernandez, Gastaldi & Hajdu View in CoL , sp. nov.
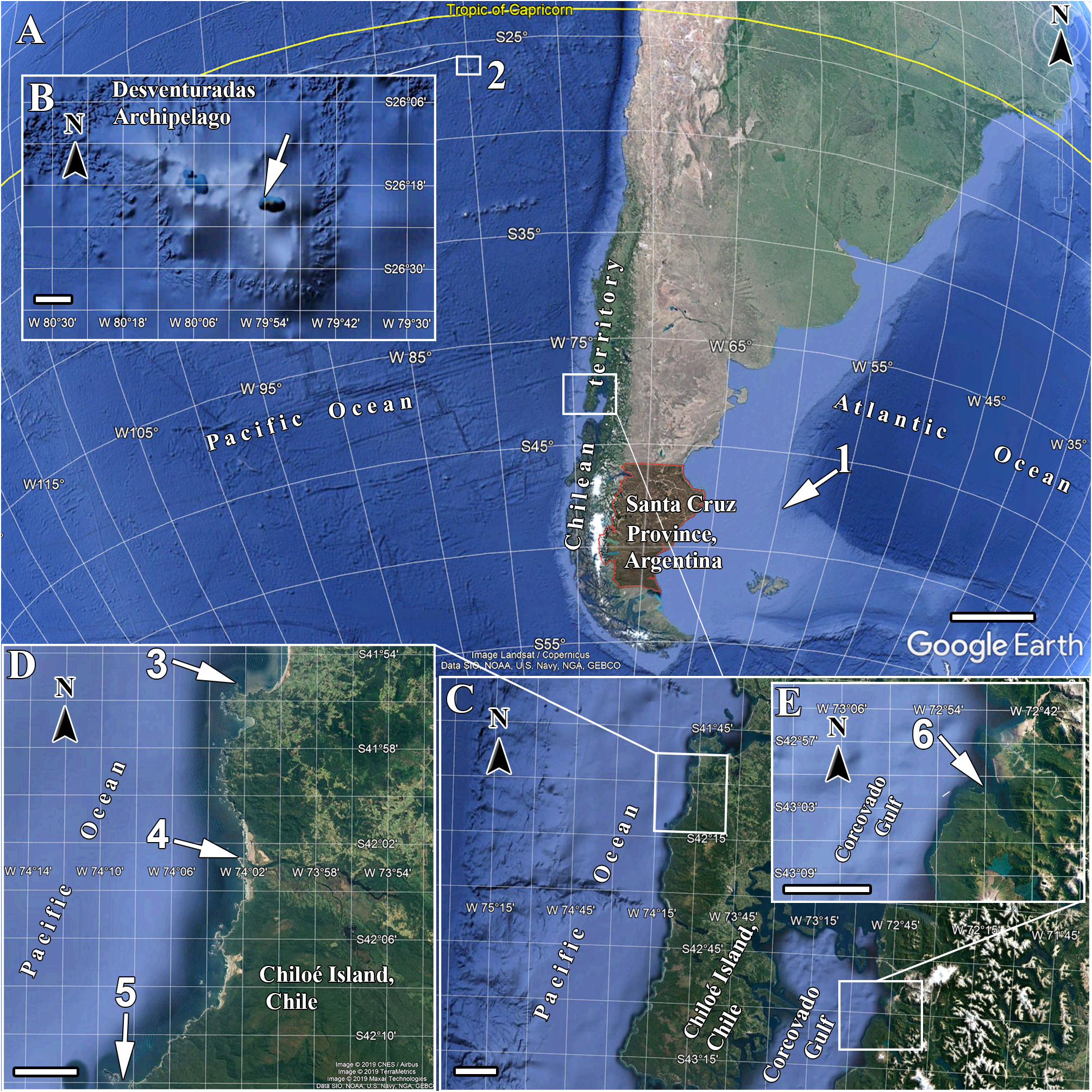
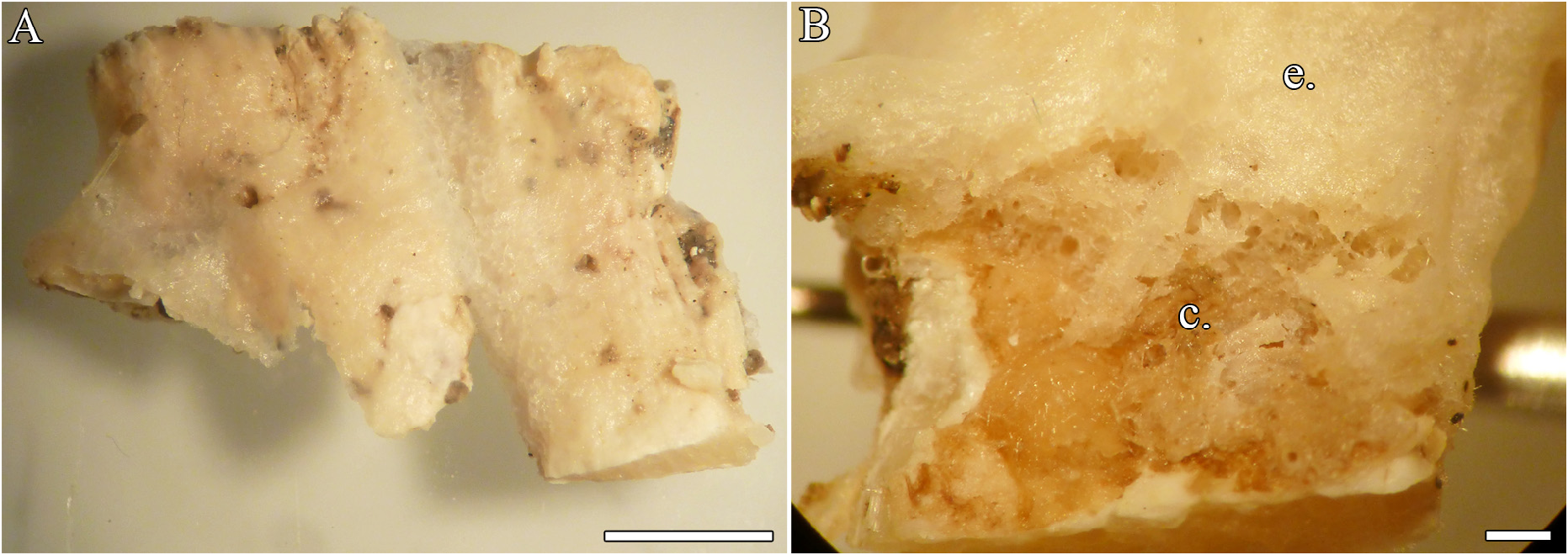
( Table 4 View TABLE 4 ; Figures 8–9 View FIGURE 8 View FIGURE 9 )
Type locality. Palvitad fjord, southern Chile .
Material examined. Holotype — IZUA-POR 172 , Rada Negra , Palvitad fjord, Corcovado Gulf, Chile (- 43.02523611 S, - 72.78222222 W), 19.7 m depth, coll. N. Reiff, 15 th March 2007. Both slides of skeleton (in low viscosity epoxy) and spicules under MNRJ 19227 View Materials . GoogleMaps
Comparative material. MNRJ 8905, 8149— Hymenanchora tenuissima ( Thiele, 1905) [= Crellomima t. sensu Willenz et al. (2009), specimens and slides].
Diagnosis. Crellomima being thin crust, with tornotes (188–255/3 – 7.2), three categories of acanthostyles (choanosomal, I. 250–285/12.5–17 µm, and II. 100–140/7.5–10 µm; ectosomal, 60–96/3.5–6.5 µm), two categories of anchorate isochelae (I. 48–60 µm and II. 12–17 µm) and sigmas (40–58 µm). So far, the only Crellomima with sigmas and two categories of anchorate isochelae.
Description. Only a small, thinly encrusting preserved fragment ( Fig. 8A View FIGURE 8 ); 1.7 (L) x 1.1 (W) x 0.3 cm (H). Surface smooth, with a thin, easily detachable membrane ( Fig. 8B View FIGURE 8 ). Oscules, pore sieves and subectosomal channels not observed. Consistency delicate, texture slightly rough. Color in life not recorded and light beige (internally slightly darker beige) in preserved specimen.
Skeleton. Plumoreticulate architecture ( Fig. 9A View FIGURE 9 ). Ectosomal region with several tornotes in vertical arrangement ( Fig. 9B View FIGURE 9 ) and in bouquets ( Fig. 9C View FIGURE 9 ), which rarely protrude to the surface (up to 50 µm high); tornotes only in the ectosomal and subectosomal regions. A continuous layer of acanthostyles (ectosomal category) at the ectosome, ca. 50 µm thick. Ectosomal acanthostyles disposed in tangential to paratangential arrangement ( Fig. 9D View FIGURE 9 ). Choanosomal region with acanthostyles (larger choanosomal category) in plumoreticulate tracts, quinated by smaller acanthostyles (smaller choanosomal category) ( Figs. 9E View FIGURE 9 ). Both categories of choanosomal acanthostyles erect on inclusions of debris and substrate too ( Fig. 9F View FIGURE 9 ). Ectosomal acanthostyles around channels at the choanosome, less usually scattered throughout the skeleton. All categories of acanthostyles (ectosomal and choanosomal) are distinguished by size (length and width) and patterns of spination. Anchorate isochelae (two categories) and sigmas (one category) scattered throughout the choanosome. In spongin, rounded subectosomal (up to 500 µm longer length) and choanosomal channels (up to 150 µm longer length). No spongin fibers throughout the skeleton. A basal spongin plate covers debris/substrates echinated by choanosomal acanthostyles ( Fig. 9F View FIGURE 9 ).
Spicules. Megascleres ( Table 4 View TABLE 4 ). Tornotes or anisotornotes ( Figs. 9G–H View FIGURE 9 ), straight to slightly sinuous, tips mucronate to lanceolate; juvenile forms slender:,188– 221.5 (19.4)–255/3– 5.8 (1.4) – 7.2 µm. Choanosomal acanthostyles I ( Fig. 9I View FIGURE 9 ), largest category of acanthostyles, usually curved, fully spined, bases blunt to slightly swollen, tips acerate; spines small (up to 5 µm high), evenly distributed, straight to slightly bent towards the bases; juvenile forms slender, with smaller, straight spines: 250– 266.7 (12.7)–285/12.5– 15 (1.6)–17 µm. Choanosomal acanthostyles II ( Fig. 9J View FIGURE 9 ), intermediate category of acanthostyles, smaller and thinner than the previuous, almost straight, fully spined; spines small (up to 3.5 µm high), evenly distributed, straight to slightly bent towards the base; juvenile forms slender, with smaller and straight spines: 100– 123.5 (13.1)–140/7. 5– 9 (0.9)–10 µm. Ectosomal acanthostyles ( Fig. 9K View FIGURE 9 ), the smallest category of acanthostyles, slightly curved to slightly sinuous, isodyametric, fully spined, bases blunt (no tyle), tips conical to acerate, acanthoxea forms not observed; spines small (up to 2.8 µm high), straight, evenly distributed; juvenile forms slender, with several smaller, thinner spines: 60– 83.8 (11.5)–96/3.5– 5.8 (1)–6.5 µm. Microscleres ( Table 4 View TABLE 4 ). Anchorate isochelae I ( Fig. 9L View FIGURE 9 ), larger, tridentate (usually) or polydentate (up to four teeth, less frequent); axis slightly curved, smooth and thicker on both extremities; alae (teeth) elongated, with slightly unguiferate tips; distance between opposite alae, ca. 1/3 of maximum chelae length; juvenile forms with slender, unguiferate alae (reduced alae): 48– 54.6 (4.8)–60 µm. Anchorate isochelae II ( Fig. 9M View FIGURE 9 ), smaller, less frequent, spatuliferous and tridentate; axis slightly curved, smooth and thicker on both extremities; alae elongated, spatuliferous and with rounded tips; very short distance between opposite alae, ca. 1/4 of maximum chelae length; juvenile forms slender and smaller: 12– 15 (2.3)–17 µm. Sigmas ( Fig. 9N View FIGURE 9 ), ‘c’ or ‘s’ shaped, smooth and sharp tips: 40– 50.5 (5.4)–58 µm.
Ecology. The specimen grew over a barnacle shell, in close association with a haplosclerid sponge; oxeas can be seen in tracts or scattered in the skeleton slide of the holotype.
Distribution. Known only from its type locality, at Palvitad fjord, southern Chile ( Fig. 1 View FIGURE 1 ).
Etymology. The name ‘sigmatifera’ is derived from sigmata (= sigma microscleres) + fera (= Latin prefix for “bearer”). The possession of sigmas microscleres is the main diagnostic character for this new species.
Remarks. Until recently Crellomima was known only from three species, all them from the Arctic Ocean; viz. C. derma Hentschel, 1929 , C. imparidans Rezvoi, 1925 (type species) and C. incrustans Hentschel, 1929 . Now, Goodwin et al. (2021) has described Crellomima mehqisinpekonuta Goodwin, Dinn, Nefedova, Nijhof, Murillo & Nozères, 2021 from northwest Atlantic. Our study presents fifth Crellomima species in worldwide.
The three Arctic species are thinly encrusting (up to 0.5 mm thick) and have a hymedesmioid skeleton ( Hentschel 1929; Rezvoi 1925). Crellomima mehqisinpekonuta is thinly encrusting (viz., author have not mentioned thickness) and has ascending columns of tornotes supported by a layer of basal acanthostyles echinating the substrate ( Goodwin et al. 2021). Crellomima sigmatifera Fernandez, Gastaldi & Hajdu , sp. nov. is also encrusting (up to 3 mm thick), but has a plumoreticulate skeleton, although, acanthostyles are echinating around embedded debris or at the base of the sponge. Furthermore, the new species is the only Crellomima species with sigmas and two categories of chelae ( Table 4 View TABLE 4 ).
Hentschel (1929) described oscules in Crellomima incrustans and Goodwin et al. (2021) observed oscules and pore fields in Crellomima mehqisinpekonuta . Other species of Crellomima had no appertures reported on their surfaces, oscula and/or pore sieves ( Hentschel 1929; Rezvoi 1925; van Soest 2002a), including the new species described here (see results above).
Willenz et al. (2009) have recorded Crellomima tenuissima ( Thiele, 1905) from the Chilean fjord region. We examined the skeleton of this material and observed acanthotornotes in longitudinal tracts throughout the choanosome as well as the same acanthose megascleres fanning out at the surface and disposed (para)tangentially at the ectosomal region. This arrangement of the acanthotornotes at the surface/ectosomal region could be equated with a tangential ectosomal crust as described by Willenz et al. (2009). However, the ectosomal crust of acanthose megascleres peculiar to Crellidae ( van Soest 2002a) is clearly differentiated from the underlying choanosomal skeleton, and not a mere consequence of the apical ends of its spiculofibres fanning out in variously oblique or tangential brushes of diactinal spicules. According to our interpretation, Willenz et al. ’s (2009) materials should be classified neither in Crellomima nor in Crellidae . This is so because these specimens lack acanthose megascleres (acanthoxeas or acanthostyles) equivalent to the acanthostyles III observed in C. sigmatifera Fernandez, Gastaldi & Hajdu , sp. nov.
Goodwin et al. (2011) have re-examined the type of Hymenanchora tenuissima [ Myxillidae Dendy, 1922 ( van Soest 2002c), originally as Hymedesmia tenuissima Thiele, 1905 ] and noted it has a plumose choanosomal skeleton, which they proposed is best classified in Myxilla (Styloptilon) Cabioch, 1968 . This skeletal architecture differs from the hymedesmioid one reported by Willenz et al. (2009), thus suggesting the latter’s materials might belong to a separate species.

| MNRJ |
Museu Nacional/Universidade Federal de Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Heteroscleromorpha |
|
Order |
|
|
Family |
|
|
Genus |