
Colobomatus stelliferi, Pombo, Maíra, Turra, Alexander, Paschoal, Fabiano & Luque, José Luis, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3925.3.8 |
|
publication LSID |
lsid:zoobank.org:pub:41704232-9307-4C1F-A7E7-3C8E197CE228 |
|
DOI |
https://doi.org/10.5281/zenodo.6101573 |
|
persistent identifier |
https://treatment.plazi.org/id/940787AF-FFCE-FFD6-FF1F-FC1B8134FF28 |
|
treatment provided by |
Plazi |
|
scientific name |
Colobomatus stelliferi |
| status |
sp. nov. |
Colobomatus stelliferi n. sp.
Figs 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4
Material examined. Holotype female from S. brasiliensis (type-host) (MZUSP-32619) collected by A. Turra, April 2004; 3 female paratypes from S. rastrifer (MZUSP-32620) collected by A. Turra, April 2004; and 3 female paratypes (MNRJ-25206) from S. stellifer collected by A. Turra, April 2004. Additional material: 22 specimens females from the Brazilian stardrum Stellifer brasiliensis (357 necropsied individuals); 47 specimens females from the rake stardrum S. rastrifer (2.852 necropsied individuals); and six specimens females from the little croaker S. stellifer (116 necropsied individuals). These additional specimens are kept in the collection of the senior author.
Type-locality. Caraguatatuba Bay, State of São Paulo, Southeastern Brazilian coast (23° 37´to 23° 44´S and 45° 24´to 45° 26´W).
Attachment site. Mandibular canals (figs. 4A, B).
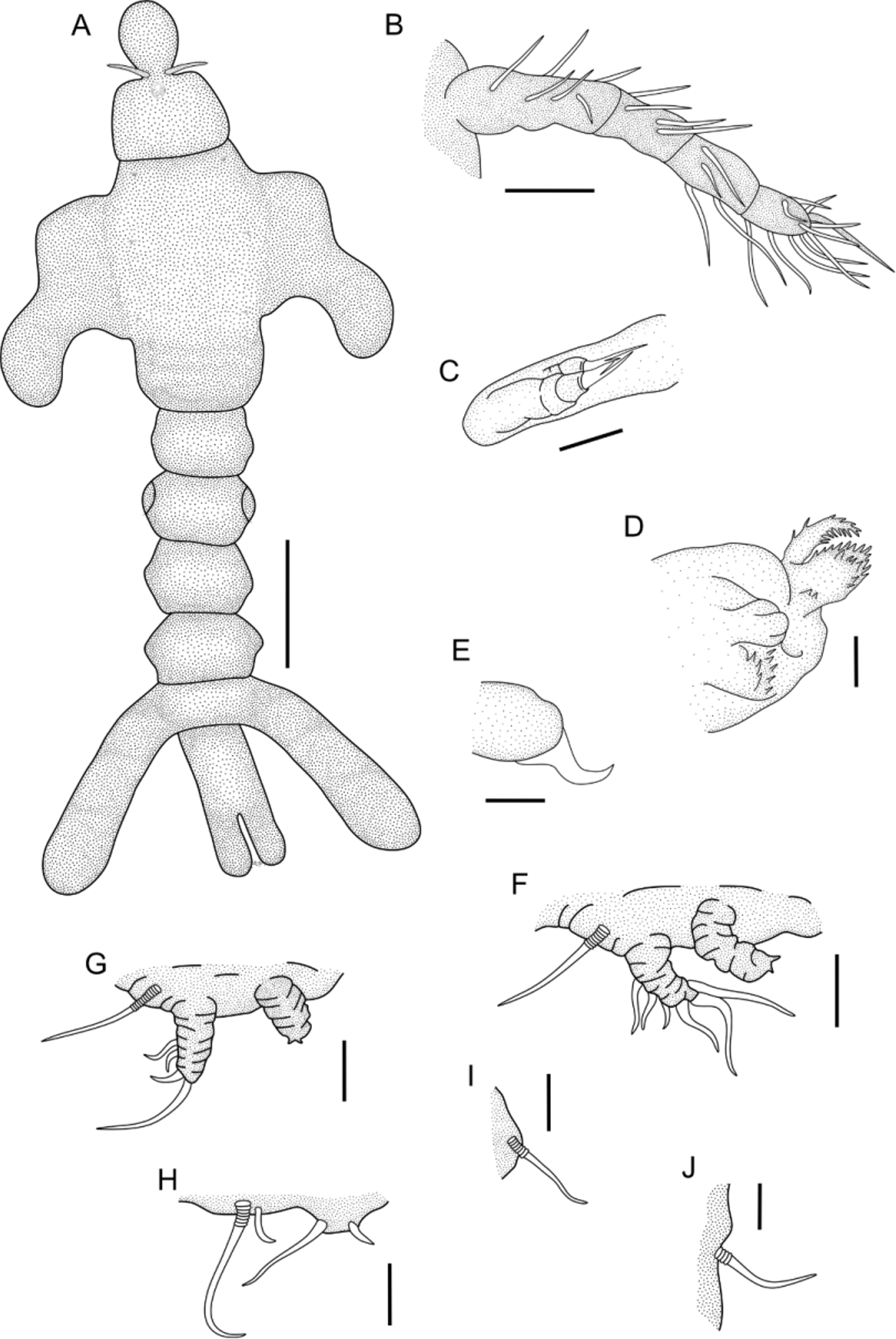
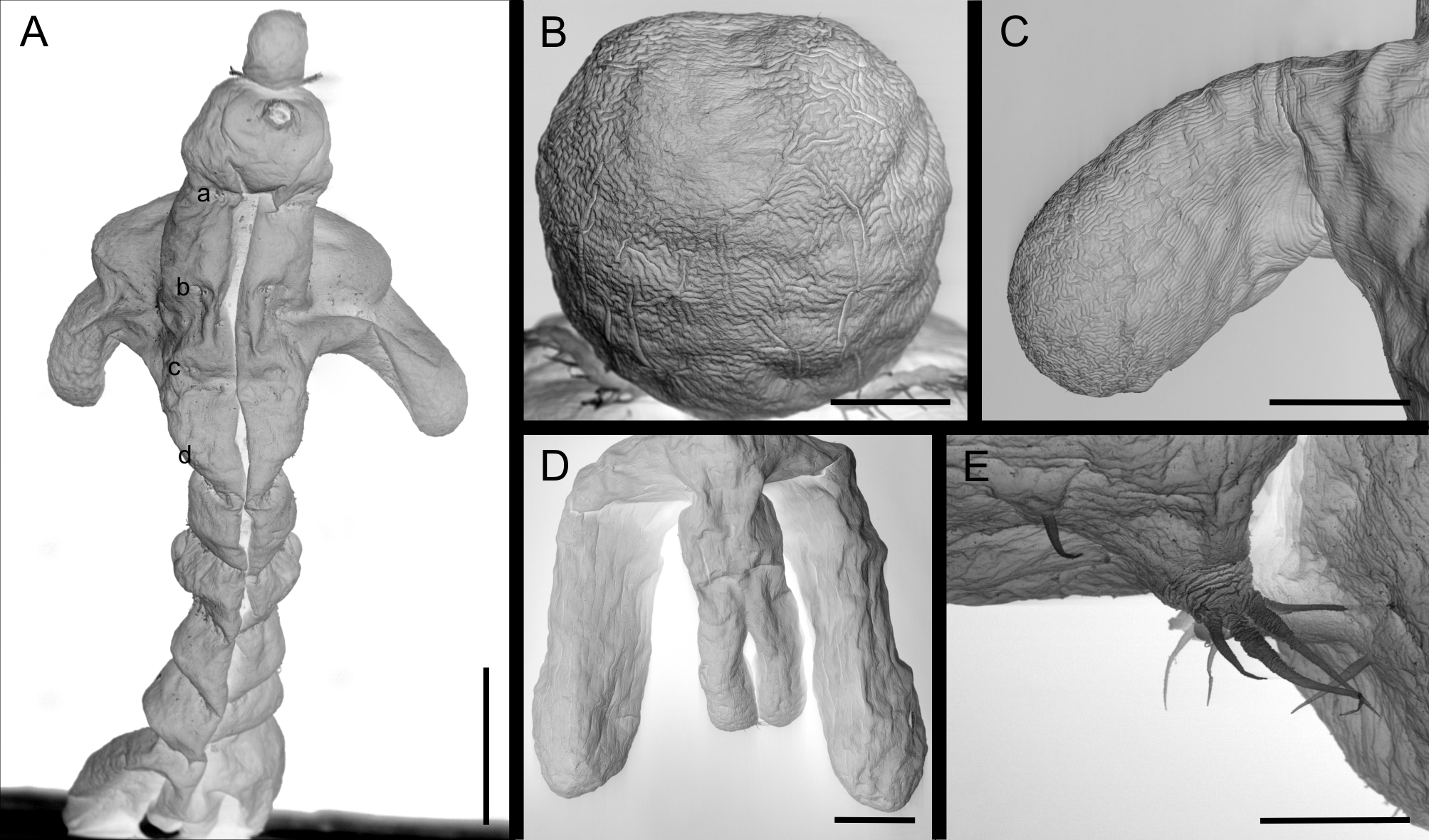
Description of adult female. Based on 10 females, all measured. Body elongate ( Figs. 1 View FIGURE 1 A, 2A, 4B), 3.31mm (2.94–3.81) long. Pre-oral area of cephalosome ( Fig. 2 View FIGURE 2 B) bearing one anterior, elongated process 264 µ m (244–308) long and 231 µ m (196–317) wide. Cephalosome trapezoidal ( Figs 1 View FIGURE 1 A, 2A), wider than long, 412 µ m (360–507) long and 563 µ m (453–710) wide, and apparently larger in posterior margin. First to fourth pedigerous somites fused, octagonal to ovoid shaped 1.01 mm (0.88–1.22) long, representing about 30% of total body length, and 826 µ m (583–990) wide, excluding dorso-lateral processes; bearing one pair of dorso-lateral processes ( Figs. 1 View FIGURE 1 A, 2A,C) located at slightly above central region. Pair of dorso-lateral processes subequally long, right process 709 µ m (521–912) long, left dorso-lateral process 708 µ m (517–920) long. First to fourth pairs of legs inserted in ventro-lateral position of fused somites. Fifth pedigerous somite shorter than wide, 293 µ m (224–350) long and 450 µ m (240–554) wide. Genital somite ( Figs. 1 View FIGURE 1 A, 3I) ovoid, 305 µ m (230–385) long, 446 µ m (340–498) wide, with pair of lateral genital orifices. Egg-sacs ( Fig. 4 View FIGURE 4 B) attached near genital orifice, 788 µ m (490–920) long (n = 4). Abdomen four-segmented. Two first abdominal somites hexagonal, first somite 253 µ m (220–291) long, 378 µ m (264–495) wide; second somite 251 µ m (210–280) long and 432 µ m (354–543) wide. Third abdominal somite ( Fig. 2 View FIGURE 2 D) with two robust lateral processes, similar to each other in size, right process 828 µ m (694–946), left process 826 µ m (698–940) long, longer than somite proper. Last abdominal somite short and gradually tapering towards caudal rami. Caudal ramus ( Figs. 1 View FIGURE 1 A, 2D) 573 µ m (457–730) long and 270 µ m (248–289) wide, with two rounded lobes bearing 4 distal caudal setae ( Fig. 2 View FIGURE 2 E).
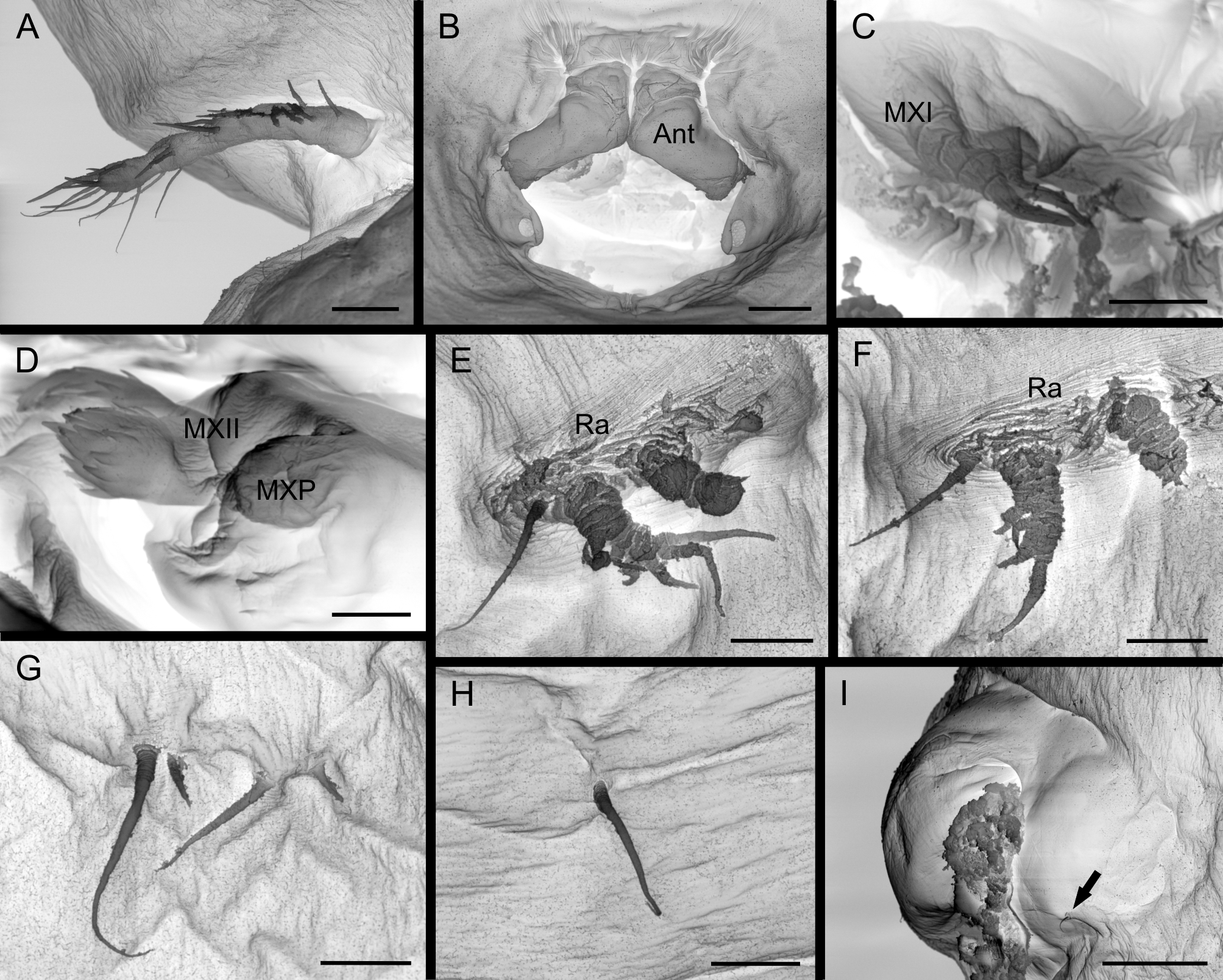
Antennule ( Figs. 1 View FIGURE 1 B, 3A) apparently four-segmented, with armature formula 6, 4, 4 and 10 + 1 aesthetasc; all antennulary setae naked. Oral area forming tube-like structure covered anteriorly by antennae and formed posteriorly by a simple, undivided labium. Antennae ( Fig. 3 View FIGURE 3 B) modified, covering anterior region of siphon-like oral area. Labrum not seen. Maxillule ( Figs. 1 View FIGURE 1 C, 3C) 1-segmented, located mid-laterally in oral area, with two apical spines. Maxilla ( Figs. 1 View FIGURE 1 D, 3D) two-segmented, basal segment with two circular rows of spinules and one apical spine; distal segment ornamented with row of spinules and two distal spinules. Maxilliped ( Figs. 1 View FIGURE 1 E, 3D) with naked basal segment and one distal spine.
Legs 1 and 2 inserted in rugose area. Leg 1 ( Figs. 1 View FIGURE 1 F, 2A, 3E) biramous, located immediately posterior to junction of cephalosome and fused somites, protopod carrying 1 lateral seta irregularly annulated at base, apparently unsegmented unarmed endopod (vestigial) and indistinctly 2-segmented exopod, armed with 3 proximal-lateral and 3 distal setae. Leg 2 ( Figs. 1 View FIGURE 1 G, 2A, 3F) biramous, located below the anterior leg, apparently in the second part of the fused somites, with protopod carrying 1 lateral seta irregularly annulated at base, apparently unsegmented unarmed endopod (vestigial) and indistinctly 2-segmented exopod, armed with 2 proximal lateral and 2 distal setae. Leg 3 ( Figs. 1 View FIGURE 1 H, 2A, 3G), located apparently in the third part of the fused somites, and reduced to 4 setae of different size, largest seta with irregularly annulated at base. Leg 4 ( Figs. 1 View FIGURE 1 I, 2A, 3H) reduced, located in the last part of the fused somites, and represented by single seta with irregularly annulated at base. Leg 5 absent. Leg 6 ( Fig. 1 View FIGURE 1 J) near genital apertures, represented by minor seta.
Male: Unknown.
Etymology. The specific name refers to type-host of the species.
Prevalence. 60% in S. brasiliensis , 16% in S. rastrifer , and 7% in S. stellifer .
Remarks. Together with the Chondracanthidae Milne Edwards, 1840 and Shiinoidae Cressey, 1975 , the Philichthyidae can be included within a group of families, characterized by the reduction of legs 4 and 5 and by the presence of 1 and 2 geniculate claws respectively on the second and third endopodal segments of the antenna in the first copepodid stage ( Boxshall & Halsey 2004). However, philichthyids can be separated based on the body shape in the adult female, it can be elongate, flattened or highly irregular with numerous processes and also philichthyid species dwell in mucous ducts of the host or form pouches in the body wall or viscera ( Boxshall & Montú 1997). According to Boxshall & Halsey (2004), this family can be divided into two groups of genera: with a body bearing at least 1 pair of lateral processes ( Colobomatoides , Colobomatus , Philichthys , Procolobomatus , Sphaerifer and Ichtyotaces) or forms without lateral processes on the body (Lephosphilus, Lernaeascus and Sarcotaces ).
Currently, the genus Colobomatus is considered to show the highest morphological diversity and is the most speciose genus with 70 species recorded, parasitizing a wide range of perciform teleosts, but also some species of Anguilliformes, Myctophiformes, Characiformes, Beloniformes, Scorpaeniformes , and one species is known from an elasmobranch species of Lamniformes ( Grabda & Linkowski 1978; West 1992; Hayward 1996; Madinabeitia et al. 2012).
Three species of Colobomatus have a single cephalic process only: C. belizensis from Haemulon spp. and Orthopristis spp. in the Neotropical Region, C. muranae (Richiardi, 1877) from Muraena helena Linnaeus in the Palearctic Region, and C. springeri Cressey, 1977 from Cryptotrema corallinum Gilbert in the Nearctic Region ( Cressey 1977; Cressey & Schotte 1983; Walter & Boxshall 2014). The above mentioned species can be easily separated from C. stelliferi n. sp. by the possession of two pairs of dorso-lateral processes on the fused pedigerous somites and the presence of lateral processes in the genital somite, whereas the new species possesses one pair of dorso-lateral processes on the fused pedigerous somites and lateral processes are absent in the genital somite.
At present only C. mackayi West, 1992 from Pomadasys striatus (Gilchrist & Thompson) in the Oriental Region has been reported to possess one pair of dorso-lateral processes on the fused pedigerous somites. C. stelliferi n. sp. is differentiated from C. mackayi by having the following features: (1) a single elongated cephalic process (two simple cephalic processes with sharp tips in C. mackayi ); (2) First to fourth pedigerous somites fused (second to fourth pedigerous somites fused in C. mackayi ); (3) absence of lateral processes in the genital somite (presence of small circular lateral processes in the genital somite of C. mackayi ); and (4) presence of lateral processes in the third abdominal somite (absent in C. mackayi ) ( West 1992).
The eight congeners that most resemble the new species are those with an absence of lateral processes in the genital somite, namely, C. arabicus Hayward, 1996 from Sillago spp. in the Palearctic Region; C. asiaticus Hayward, 1996 from Sillago spp. in the Oriental Region; C. bergyltae Hesse, 1873 from Labrus bergylta Ascanius in the Palearctic Region; C. caribbei Cressey & Schotte, 1983 from Anisotremus surinamensis (Bloch) in the Neotropical Region; C. cribbi West, 1992 from Scolopsis taenioptera (Cuvier) in the Australian Region; C. pagelli (Richiardi, 1877) from Pagellus spp. and Lithognathus mormyrus (Linnaeus) in the Palearctic Region; C. quadrifarius Cressey & Schotte, 1983 from Anisotremus spp., Haemulon spp. and Orthopristis spp. in the Nearctic and Neotropical Regions; and C. sieboldi (Richiardi, 1877) from Boops boops (Linnaeus) in the Palearctic Region ( Cressey & Schotte 1983; West 1992; Hayward 1996; Walter & Boxshall 2014). Colobomatus stelliferi n. sp. can be readily distinguished from all eight above mentioned congeners by the presence of a single cephalic process in the cephalosome and one pair of dorso-lateral processes on the fused pedigerous somites. Each of the other eight species possesses two cephalic processes in the cephalosome and two pairs of dorso-lateral processes on the fused pedigerous somites ( Yamaguti 1963; Cressey & Schotte 1983; West 1992; Hayward 1996).
The majority of species of Colobomatus are parasites of perciforms fishes and three species have been recorded from sciaenid fishes. These species are C. orientalis Kim & Moon, 2013 from Johnius grypotus (Richardson) ; C. sciaenae (Richiardi, 1876) from Argyrosomus regius (Asso) and Sciaena umbra Linnaeus , both in the Palearctic Region; and Colobomatus sp. from Micropogonias furnieri in the Neotropical Region. The findings of this study reaffirming the status of sciaenid fishes as hosts for the genus Colobomatus and extend the host range to include members of genus Stellifer . However the family Sciaenidae are distributed in the Atlantic, Indian and Pacific oceans and possess an elevated number of species, approximately 283 spp. distributed in 66 genera, which probably indicate gaps in the real diversity of parasite copepods of the family Philichthyidae in these host group.
According to Grabda (1991) species of the genus Colobomatus possess highly specific by a single host species. However, based on the limited Grabda’s data, Hayward (1996) proposed that most species are specific to genera or families host rather than to a single species. Recently, Madinabeitia et al. (2013) commented on the host-specificity of C. pupa Izawa, 1974 by members of the genus Parupeneus (i.e. P. spilurus (Bleeker) , P. ciliatus (Lacépède) and P. multifasciatus (Quoy & Gaimard)) . Consequently, we support Hayward’s hypothesis of Colobomatus spp.
exhibiting a high host-specificity at the genus level, C. stelliferi n. sp. is probably specific to members of the genus Stellifer (i.e. S. brasiliensis , S. rastrifer and S. stellifer ).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |